

Психиатрия Всемирная психиатрия
№02 2020
Что такое резильентность: аффилиативный нейробиологический подход №02 2020
Номера страниц в выпуске:132-150
Резюме
Резильентность (жизнестойкость, психологическая устойчивость – прим. редактора) – ключевая проблема клинической науки и практики – все еще не имеет четкой концепции, которая объединила бы ее эволюционные и специфичные для человека свойства, при этом не ограничиваясь изучением исключительно физиологии страха, включала бы в себя целостный подход с позиций общего развития и, что самое важное, чтобы ее определение не было отрицательным (т. е. отсутствие симптомов после травмы). Опираясь на особенности первоначального состояния млекопитающих, чей мозг созревает как в процессе внутриутробного развития, так и в процессе воспитания, мы утверждаем, что системы и процессы, которые участвуют в настройке мозга на социальную экологию и адаптации к ее трудностям, характеризуют концепцию резильентности. К ней относятся система окситоцина, аффилиативный мозг и биоповеденческаяя синхронность, которые характеризуются значительной гибкостью в филогенезе и онтогенезе. Выделены три основных принципа резильентности: пластичность, социальность и осмысленность. Механизмы социального взаимодействия, с помощью которых согласованные действия обеспечивают разнообразие, выносливость и адаптацию, описаны в эволюции животных. Биоповеденческая синхронность человека постепенно развивается от «сонастроенного» поведения матери в послеродовом периоде к отношениям «взрослый–взрослый», наполненным эмпатией, взаимопониманием и близостью, и простирается от взаимоотношений матери и ребенка на другие аффилиативные связи на протяжении всей жизни, намечая фундаментальную траекторию в развитии резильентности. Результаты исследования трех когорт высокого риска, каждая из которых характеризуется явными нарушениями детско-материнских взаимоотношений (недоношенность, материнская депрессия и перенесенный в раннем возрасте стресс/психотравма), с периодом катамнестического наблюдения от рождения до подросткового возраста / ранней молодости, демонстрируют, как нейробиологические компоненты аффилиации обеспечивают резильентность и уникальным образом формируют социальный мозг.
Ключевые слова: резильентность, система окситоцина, аффилиативный мозг, биоповеденческая синхронность, детско-материнские отношения, нейробиология аффилиации, социальность, пластичность, смысл.
Резильентность (жизнестойкость, психологическая устойчивость – прим. редактора) – ключевая проблема клинической науки и практики – все еще не имеет четкой концепции, которая объединила бы ее эволюционные и специфичные для человека свойства, при этом не ограничиваясь изучением исключительно физиологии страха, включала бы в себя целостный подход с позиций общего развития и, что самое важное, чтобы ее определение не было отрицательным (т. е. отсутствие симптомов после травмы). Опираясь на особенности первоначального состояния млекопитающих, чей мозг созревает как в процессе внутриутробного развития, так и в процессе воспитания, мы утверждаем, что системы и процессы, которые участвуют в настройке мозга на социальную экологию и адаптации к ее трудностям, характеризуют концепцию резильентности. К ней относятся система окситоцина, аффилиативный мозг и биоповеденческаяя синхронность, которые характеризуются значительной гибкостью в филогенезе и онтогенезе. Выделены три основных принципа резильентности: пластичность, социальность и осмысленность. Механизмы социального взаимодействия, с помощью которых согласованные действия обеспечивают разнообразие, выносливость и адаптацию, описаны в эволюции животных. Биоповеденческая синхронность человека постепенно развивается от «сонастроенного» поведения матери в послеродовом периоде к отношениям «взрослый–взрослый», наполненным эмпатией, взаимопониманием и близостью, и простирается от взаимоотношений матери и ребенка на другие аффилиативные связи на протяжении всей жизни, намечая фундаментальную траекторию в развитии резильентности. Результаты исследования трех когорт высокого риска, каждая из которых характеризуется явными нарушениями детско-материнских взаимоотношений (недоношенность, материнская депрессия и перенесенный в раннем возрасте стресс/психотравма), с периодом катамнестического наблюдения от рождения до подросткового возраста / ранней молодости, демонстрируют, как нейробиологические компоненты аффилиации обеспечивают резильентность и уникальным образом формируют социальный мозг.
Ключевые слова: резильентность, система окситоцина, аффилиативный мозг, биоповеденческая синхронность, детско-материнские отношения, нейробиология аффилиации, социальность, пластичность, смысл.
Резильентность (жизнестойкость, психологическая устойчивость – прим. редактора), обычно определяемая как способность преодолевать трудности, добиваясь успеха1-3, вероятно, является конечной целью зрелой личности и наиболее важной целью лечения и профилактики. Стать человеком, способным мужественно и настойчиво справляться с жизненными трудностями, сохранять позитивный взгляд на мир в трудных условиях, получать удовольствие от общения в узком и широком кругах, проявлять эмпатию и сострадать несчастьям других, сохранять усердие и активную жизненную позицию на пути к собственным долгосрочным целям, жить жизнью, наполненной творчеством, энергией и смыслом, и не иметь изнуряющих симптомов, несмотря на пережитые невзгоды или текущие травмирующие обстоятельства, является исключительным человеческим достижением и основной целью психотерапии со времен Фрейда. Тем удивительнее, что несмотря на десятилетия исследований комплексная биоповеденческая концепция резильентности еще не сформулирована.
Текущие исследования резильентности обычно фокусируются на нейробиологии стресса и страха или основаны на эпидемиологических и клинических исследованиях последствий психотравмы. В обоих случаях резильентность понимается как «отсутствие симптомов» или «сохранение психического здоровья» после перенесенных несчастий или травм4. Междисциплинарная группа5, подчеркивая необходимость смещения акцента в области психического здоровья с психопатологии на резильентность и указывая на огромное экономическое бремя и субъективные страдания, вызываемые расстройствами, связанными со стрессом, пришла к выводу, что резильентность может быть определена только постфактум травмы, когда у части людей появляются симптомы, а у части – нет.
С научной точки зрения такая позиция является проблемной. Без четкой концепции невозможно накапливать эмпирические данные и сведения о проведенном лечении.
В частности, очень важно определить, относятся ли к резильентности процессы, которые приводят к ухудшению состояния после физической или психической травмы, или те, которые однозначно способствуют увеличению силы и выносливости6,7.
Позитивная психология выделяет резильентность как ключевую составляющую благополучия8, в ее рамках запущен известный тренинг резильентности в армии США9. Также некоторые аспекты резильентности отражены в работах психоаналитиков-постфрейдистов, которые подчеркивали, что «я» является функциональным, развивающимся, существующим не изолированно, но во взаимодействии с социальной средой, в частности в трудах Sullivan10, Fromm11 и Erickson12, в работе Maslow13 о самореализации и в понятиях гуманистической психологии14. Тем не менее, эти авторы говорили не о резильентности как таковой, а о личностном росте, не интегрировали результаты систематических исследований и данные нейробиологических открытий в свои модели и концепции или даже отрицали релевантность каких-либо нейронаучных данных15,16. Специфичная для человека модель резильентности, которая, с одной стороны, учитывает его внутреннюю реальность и высшие способности, а с другой стороны, опирается на эволюционные модели и включает в свою основную концепцию результаты нейронаучных открытий, так и не была построена.
Две основные проблемы способны еще больше усложнить построение комплексной биоповеденческой модели резильентности человека. Во-первых, большинство современных усилий, направленных на понимание нейробиологических основ психических расстройств в области психиатрии, как правило, сосредоточены на особенностях, которые можно легко проверить в сравнении с другими видами. Это привело к тому, что практически все внимание сфокусировано на нейробиологии страха – нейронных, эндокринных, генетических и молекулярных процессах, которые поддерживают реакцию страха и позволяют справляться со стрессом4,17-21. Соответственно, в исследованиях часто используются межвидовые парадигмы изучения стресса, в частности механизмов, обуславливающих страх, что привело к взгляду на резильентность преимущественно с этой точки зрения22,23.
Во-вторых, не уделяется должное внимание развитию как ключевому компоненту психического здоровья, в частности резильентности, несмотря на тот факт, что все «я»-модели, по сути, являются моделями развития (то есть описывают стадийное развитие от незрелых форм к зрелым). С хорошей резильентностью не только рождаются, ее (что важно) можно воспитать. Было высказано предположение24, что для изучения резильентности мы должны обследовать детей с младенчества в течение продолжительного периода времени, чтобы выявить возрастные биологические, поведенческие и социальные маркеры, способствующие формированию у них жизнестойкости. Однако такие лонгитудинальные исследования крайне редки.
Эти две проблемы привели к довольно ограниченному, одностороннему взгляду на резильентность. Когда в дискуссии по поводу резильентности группе ведущих исследователей25 был задан вопрос «что должно быть пересмотрено», все они указали на узкое понимание резильентности, сведение ее до физиологии страха и нейробиологии стресса в теоретических и практических исследованиях как на главную проблему, заслуживающую внимания.
Новая концепция резильентности должна основываться на эволюционных механизмах, позволять проводить основательные межвидовые исследования и создавать условия для тщательного сбора данных для выявления ее специфических особенностей на разных стадиях развития, в разных условиях. С точки зрения науки чрезвычайно важно, чтобы результаты были проверяемыми (то есть открытыми для проверки уровня доказательности и фальсификации).
В дальнейшем мы предлагаем модель резильентности, основанную на нейробиологии аффилиации и предлагающую биоповеденческую эволюционную концепцию, учитывающую аспекты развития, которая не ограничена нейробиологией страха или стремлением к счастью. Наша модель учитывает фундаментальную особенность млекопитающих, чей мозг созревает как в процессе внутриутробного развития, так и в процессе воспитания, и утверждает, что созревание всех нейробиологических процессов, которые способствуют формированию резильентности, обеспечивается физиологией материнского тела и типичным для вида родительским поведением.
Кроме того, модель утверждает, что любое понимание резильентности должно учитывать первоначальную зависимость младенца от матери и огромное влияние, которое эта зависимость оказывает на структуру и функционирование мозга. Детеныши млекопитающих рождаются с незрелым мозгом, их мозг формируется под влиянием зрелой матери через физическую близость, лактацию и совокупность специфических для вида, способствующих хорошей адаптации способов ухода за детенышем, которые обеспечивают внешнюю регуляцию в системе «мать – ребенок» уникальным для системы образом26,27.
Такая внешняя регуляция незрелого мозга зрелым определяет основной механизм развития мозга у млекопитающих и обеспечивает адаптацию нейробиологической и поведенческой систем ребенка к жизни в рамках социальной экологии и ее уникальных особенностей28,29. Мы утверждаем, что процессы адаптации мозга ребенка к жизни в пределах экологической ниши и тем явным трудностям, с которыми он сталкивается, и есть сама суть резильентности, именно эти процессы определяют, что такое резильентность, и они должны стать центром внимания научных исследований.
ОСНОВНЫЕ НЕЙРОБИОЛОГИЧЕСКИЕ КОМПОНЕНТЫ АФФИЛИАЦИИ
Наша модель опирается на три основных нейробиологических компонента аффилиации: окситоциновая система, аффилиативный мозг и биоповеденческая синхронность.
Окситоциновая система
Древняя окситоциновая система, появившаяся примерно 500 миллионов лет назад, обеспечивала реакции организма на экологические проблемы, участвуя в регулировании основных жизненных функций, таких как сохранение водного и энергетического баланса, терморегуляцию в филогенетическом масштабе31,32. Таким образом, первоначально окситоцин обеспечивал выносливость, адаптацию организма к условиям внешней среды и его резильентность.
В процессе эволюции у млекопитающих окситоцин стал играть ключевую роль в родах и лактации. Тогда для млекопитающих связь между матерью и детенышем стала ключевым условием для созревания систем, которые обеспечивали совладание со стрессом32. Жизненно важные функции больше не развиваются в контексте группы, как у рыб или муравьев, а, в интимных рамках «кормящей диады», благодаря особым структурам материнского тела.
У млекопитающих окситоциновая система стала основой, обеспечивающей механизмы резильентности и аффилиации, посредством которых устойчивость, пластичность и толерантность к трудностям внешнего мира достигаются за счет социального взаимодействия, которое связывает одну особь с человеческим культурным сообществом29,33.
В целом роль окситоцина для резильентности определяется тремя компонентами, связанными с его вовлечением в нейропластичность, социальность и иммунитет.
Окситоцин задействован в процессах нейропластичности на молекулярном, клеточном уровнях и уровне нейрональной сети34-36. Нейроны могут вырабатывать окситоцин совместно с различными нейротрансмиттерами, в том числе дофамином, серотонином и опиоидами. Окситоцин вырабатывается широким спектром нейрональных клеток, таких как ГАМКергические интернейроны, глутаматергические пирамидальные клетки и другие пептидергические клетки34,37,38. Окситоцин объединяет центральные и периферические отделы нервной системы, включает массивные эпигенетические данные и, в частности, связан с переживаниями привязанности39,40. Он повышает пластичность в сети гиппокампа, повышая привлекательность объекта привязанности41, а приобретенный опыт привязанности влияет на доступность окситоциновых рецепторов42.
Пульсирующий характер высвобождения окситоцина имеет особое значение для нейропластичности, благодаря которой формируются нейробиологические системы взаимодействия с окружающей средой43. Пульсирующее высвобождение окситоцина координирует рождение при благоприятных условиях окружающей среды, знаменуя первое в жизни человека взаимодействие мозга и окружающей среды44. Всплеск окситоцина во время рождения вызывает передачу сигналов посредством гамма-аминомасляной кислоты (ГАМК), что приводит к переходу от возбуждения к торможению, тем самым координирует переход нейронов гиппокампа плода от функционирования в пренатальном к функционированию в постнатальном периоде45,46, поддерживая баланс между возбуждением и торможением на протяжении всей жизни. Оптимальный баланс возбуждения и торможения имеет решающее значение для адаптивного функционирования и обеспечивает эффект «сензитивного периода», который имеет решающее значение для устойчивости всех живых организмов47.
Окситоцин играет ключевую роль в формировании социальности. Нейронные сети, обеспечивающие формирование привязанности и связей, развивались из-за чувствительности окситоцина к повторяющимся элементам в окружающей среде, благодаря чему мать и окружение становились ценными стимулами48-50. Окситоцин в лимбических структурах побуждает младенцев предпочитать сигналы, связанные с матерью, что приводит к формированию специфической диадной привязанности51,52. В течение первых дней после рождения рецепторы окситоцина становятся ассоциированными со специфическими социальными сигналами через связи системы окситоцина с дофаминной системой вознаграждения53-55, ольфакторно-амигдалярными путями56,57, сенсорной корой54 и резким повышением отношения «сигнал/шум» пирамидальных клеток гиппокампа58. Они программируют социальное восприятие, предпочтения и память и связывают их с объектом привязанности.
Окситоцин способствует интеграции людей в социальные группы59,60. В ходе эволюции он участвовал в обеспечении социальных функций: ритмичных движениях при ухаживании у нематод61, социальных процессах у червей62, формировании пар и стай у птиц63, исключительной связи у стадных животных64 и принадлежности к группе у грызунов65, приматов66 и людей29,67. Эволюционные ограничения привели к тому, что эта гибкая, изменчивая в зависимости от внешних условий система заставляла молодых особей находиться в связи со своими родителями, функционировать в рамках их социальной экологии и включаться в социальную структуру своего вида64. Примечательно, что большая социальная поддержка и чувство принадлежности к социальной группе неоднократно ассоциировались с лучшей резильентностью68-70.
Система окситоцина у ребенка формируется в процессе ухода за ним. Исследования на животных показывают, что материнское поведение программирует доступность окситоциновых рецепторов в мозге71, а лонгитудинальные исследования на людях показывают, что окситоцин в периферической нервной системы программируется повторяющимися проявлениями заботы в детстве72-74. Окситоцин вызывает физиологическое состояние покоя, которое позволяет без страха проявлять себя в социуме и стимулирует стремление к социальному контакту через связи с дофамином в нейронах стриатума75-77. Это уникальное состояние обеспечивает основу для чувства безопасности человека, на котором может формироваться резильентность.
Наконец, окситоцин играет важную роль в функционировании иммунной системы. Исследования на людях показали связь между окситоцином и иммунными биомаркерами78,79. В клеточной культуре окситоцин снижает окислительный стресс и секрецию интерлейкина-6 (IL-6) из стимулированных макрофагов80. In vivo он снижает уровень воспалительных цитокинов, IL-6 и фактора некроза опухоли (TNF)-α81. Во время образования связей, в том числе в период становления родителем и влюбленности, и окситоцин, и IL-6, иммунный биомаркер, повышают свою активность82, а окситоцин участвует в более быстром заживлении ран83. Недавно было установлено, что кишечная бактерия, продуцирующая окситоцин (Lactobacillus reuteri), играет роль в резильентности, совладании со стрессом и более быстром заживлении ран у хозяина, что указывает не только на наличие дополнительной оси «мозг–кишечник» в выработке окситоцина, но и на значение связи «микробиом–человек» для формирования резильентности84.
Аффилиативный мозг
Термин «аффилиативный мозг» обозначает сеть взаимосвязанных структур, которые позволяют людям формировать и поддерживать близкие отношения85.
Аффилиативный мозг человека, имеющий происхождение от материнского мозга грызунов, расширился и включил в себя несколько корковых сетей высшего порядка, которые обеспечивают бессознательную мотивацию и когнитивные аспекты воспитания человека30,86. Эта глобальная нейрональная сеть, целью которой изначально являлось обеспечение ухода и связи младенца с матерью в ранние сензитивные периоды, в дальнейшем преобразовалась для поддержания социальной связи с другими людьми: любовниками, близкими друзьями и собратьями30,85.
Исследования материнского мозга на животных моделях, предпринятые в 1950-х гг., описывают основополагающую роль медиальной преоптической области гипоталамуса в подкорковой сети, которая позволяет матерям млекопитающих (и отцам у двуполовых видов) заботиться о детенышах87. Под действием окситоцина во время беременности и родов медиальная преоптическая область направляет импульсы в миндалину, что вызывает повышение осторожности у матери для обеспечения безопасности ребенка, и в вентральную область покрышки, что активирует материнскую систему вознаграждения от стимулов младенца, повышает чувствительность лимбической сети, поддерживающей материнское поведение (также включая прилежащее ядро, боковую перегородку, вентральный паллидум, ядро ложа конечной полоски и бледное ядро).
У людей эта подкорковая сеть расширилась и включает сети более высокого порядка, которые ответственны за эмпатию, подражание, ментализацию и регулирование эмоций, формируя глобальную сеть, обеспечивающую привязанность30. У 3–5% видов млекопитающих, у которых уход обеспечивают оба родителя, такая же система лежит в основе родительского поведения отца. Однако недавние исследования показали, что у матерей и отцов родительское поведение обеспечивают разные популяции нейронов88, и за отцовское, и материнское поведение отвечает одна и та же сеть, но путь к отцовству является более «когнитивным» и опирается на конкретные действия по уходу за детьми87,89.
Окситоцин играет важную роль в настройке и функционировании аффилиативного мозга. Социальное поведение людей регулируется сетью обеспечения ухода, которая содержит множество рецепторов окситоцина90. Окситоцин вызывает долгосрочную депрессию в миндалине91, чтобы ослабить ее ответ на отрицательные социальные стимулы, таким образом укрепляя сетевое соединение и обеспечивая специфичность ответа на социальные стимулы92,93.
После ослабления социального избегания окситоцин усиливает мотивацию к формированию социальных связей через перекрестные взаимодействия с дофаминовыми рецепторами в стриатуме, в частности в прилежащем ядре. Дофамин в прилежащем ядре обеспечивает целенаправленное поведение, связанное с вознаграждением, подавляет активность ГАМКергических (ингибирующих) нейронов94-97, что позволяет активировать глутаматные (возбуждающие) пути, что в свою очередь приводит к энергичным, активным, целенаправленным действиям98,99.
Оболочка прилежащего ядра содержит рецепторы окситоцина, которые образуют гетеромеры (нейроны, вырабатывающие и окситоцин, и дофамин100), и это позволяет дофаминовым нейронам, основным назначением которых является идентификация сенсорно-моторного вознаграждения, кодировать временные паттерны социального взаимодействия49,101. Это позволяет мозгу опознавать социального партнера, кодировать специфические для этой связи паттерны и получать вознаграждение от социальной синхронности96,101.
Тесные перекрестные связи «окситоцин–дофамин» позволяют гибко встраивать новую связь в личность102 и формировать сенсорно-моторные воспоминания о переживаниях привязанности103. Таким образом, в то время как дофамин обеспечивает мотивацию и активность, окситоцин обеспечивает спокойствие, необходимое для образования связи.
Эта мозговая сеть обеспечивает не только родительское поведение, но и нейронную поддержку для формирования других аффилиативных связей на протяжении всей жизни; отсюда и термин «аффилиативный мозг». В исследованиях животных104 и человека85 показано, что родительский мозг млекопитающих также участвует в поддержании парных связей у моногамных млекопитающих77, а также романтической привязанности и близких дружеских отношений у людей85. Эта аффилиативная сеть развивается в мозгу младенца в ранние сензитивные периоды благодаря целенаправленной заботе и позволяет ребенку формировать близкие отношения, влюбляться, становиться членом социальных групп от спортивной команды до наций и в конечном итоге, воспитывать своих детей.
Еще Darwin105 отмечал, что эволюционные изменения происходят во взаимодействии «родитель–детеныш», а присущая им пластичность обеспечивает появление новых форм поведения, которые со временем изменяют экспрессию генов. Соответственно, наша модель, в основе которой лежит взаимодействие «родитель–ребенок», показывает, как аффилиативный мозг использует присущую ему пластичность для формирования резильентности, выносливости и перенастройки.
Аффилиативный мозг обеспечивает резильентность несколькими способами. Оптимальная активация этой сети позволяет людям формировать и поддерживать социальные связи на протяжении всей жизни, справляться со стрессом с помощью отношений и благодаря перекрестным связям окситоцина и дофамина получать наибольшее вознаграждение от этих связей, а не из несоциальных источников (например, употребление наркотиков). Действительно, при зависимостях обнаружены нарушения взаимоотношений окситоцина и дофамина, что приводит к тому, что вознаграждение разъединяется с его социальными источниками, при этом выявлены нарушения как системы окситоцина106, так и нейропластичности107,108.
Мозг родителя формирует социальные способности ребенка. Мы обнаружили, что активность мозга родителя в младенчестве позволяла прогнозировать способность к регуляции эмоций, совладанию со стрессом и формирование симптомов у ребенка в течение первых семи лет жизни109-111. Вместе с тем заботливое и отзывчивое воспитание в перспективе обеспечивало формирование аффилиативного мозга в подростковом возрасте112,113. Наконец, большая ассоциативная кора человеческого мозга позволяет обрести смысл через любовь к абстрактным идеям, таким как родина или Бог, и распространять привязанность на других людей, домашних животных, флору и фауну Земли, и все это обеспечивает одна и та же сеть85.
Биоповеденческая синхронность
Биоповеденческая синхронность является основным механизмом, обеспечивающим социальность и аффилиацию у человека. Она определяется как «координация биологических и поведенческих сигналов между социальными партнерами в момент социального контакта» и описывает механизм, с помощью которого зрелый мозг родителя внешне регулирует незрелый мозг младенца и настраивает его на социальную жизнь29,114,115.
Биоповеденческая синхронность создает шаблон для координации биологического с социальным и психическим, делает возможным слияние автономного «я» с автономным «другим» и объединение моментов межличностного соответствия с моментами несоответствия, моментов разлуки и встречи – все в рамках безопасного диалога.
В многочисленных исследованиях, охватывающих период от младенчества до взрослой жизни среди широкого круга здоровых и групп населения высокого риска в различных культурах, мы показали, что эти «драгоценные социальные моменты», когда родитель и ребенок координируют свое невербальное поведение, представляют собой моменты биологической координации. Например, только во время этих эпизодов наблюдалась синхронность между сердечными ритмами матери и младенца116, координированный выброс окситоцина117 и синхронизация социального мозга обоих118.
Синхронность сочетается с лучшим совладанием со стрессом73, более высокой дыхательной синусовой аритмией119 и лучшим функционированием иммуннитета120, открывая механизм, посредством которого скоординированное социальное поведение снижает стресс и улучшает резильентность.
Связь поведенческой и биологической синхронности берет свое начало в утробе матери121, когда биологические ритмы младенца включаются в социальный диалог, который превращает биологическое в отношенческое, а индивидуальное – в межличностное. Паттерны невербальной синхронности проявляются в диадических отношениях постоянно, расширяясь в символической и межличностной сложности122, и, как предполагает теория динамических систем123, такое расширение репертуара с опорой на первичный опыт отражает механизм резильентности. Примечательно, что все формы физиологической синхронности (нейронная, эндокринная и автономная) встроены в поведенческую координацию, что подтверждает нашу основную гипотезу о том, что поведенческая синхронность формирует физиологическую связь и что резильентность базируется на поведении29,124,125.
Биоповеденческая синхронность, наблюдаемая в первые месяцы жизни, представляет собой крайне важный опыт, полученный в сензитивный период, который позволяет прогнозировать множество компонентов, связанных с резильентностью от рождения до юности, в том числе регуляцию эмоций, символическую компетентность, способность справляться со стрессом, выраженность экстернализированных и интернализированных симптомов, а также социальные развитие мозга30,114,126,127.
В процессе развития сопоставление ребенком невербальных аффективных реакций превращается в обоюдный обмен, который порождает, как брошенный камень порождает круги на воде, растущие символические, лингвистические и социальные компетенции ребенка, развивается эмпатия, мировоззрение и близость, все они построены на ритмичном невербальном ядре и служат резильентности (см. ниже). Это перекликается с представлениями Maslow13 о том, что «я» включает в себя как то, чем является человек, так и то, кем он может стать. В то же время, будучи специфическим для человека механизмом, который развивается в течение всей человеческой жизни, биоповеденческая синхронность опирается на длинную эволюционную линию необходимых для выживания социальных механизмов млекопитающих и других эусоциальных (гиперсоциальных) видов, обеспечивающих выносливость и резильентность.
В ходе эволюции, от бактерий до человека, синхронность основывается на процессах, которые связывают два организма (или две сущности) в единую биологию. Последние достижения в области квантовой физики позволяют предположить, что такое взаимодействие началось еще до появления жизни, что можно увидеть в феномене «квантовой запутанности», при котором связь частиц во времени и пространстве, объединяющая две единицы вместе, придает их объединению неизмеримую силу и стойкость.
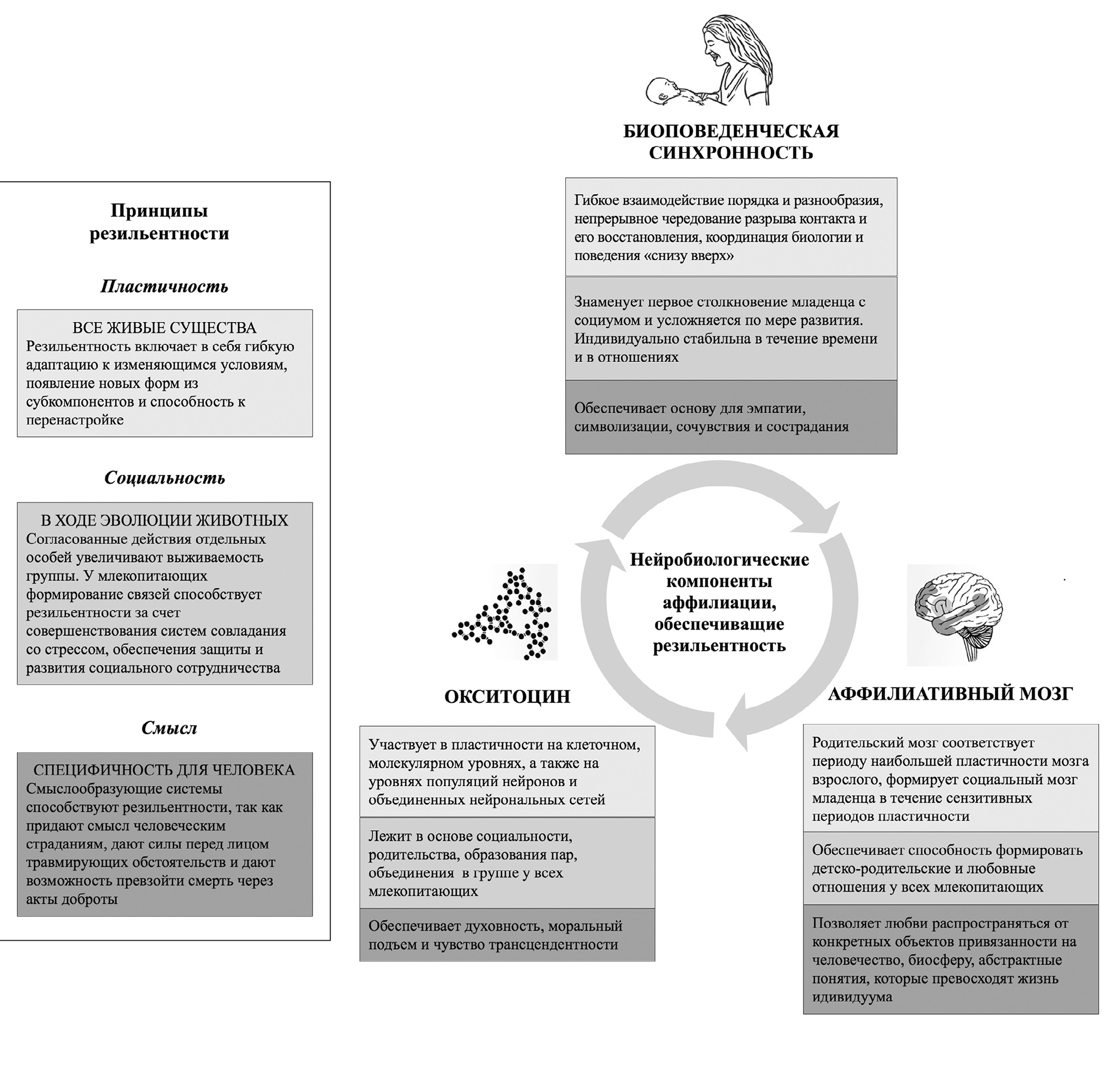
ТРИ ПРИНЦИПА РЕЗИЛЬЕНТНОСТИ
Учитывая вышеупомянутые основы аффилиативной нейробиологии (окситоциновая система, аффилиативный мозг и биоповеденческая синхронность), наша модель выделяет три принципа, определяющих, что такое резильентность. Несмотря на то, что все три элемента необходимы для формирования жизнестойкости человека, они существуют в различных комбинациях у разных людей в разных культурах и выражаются по-разному в зависимости от возраста и стадии (Рисунок 1).
Резильентность включает в себя пластичность
Прежде всего, резильентность включает в себя механизмы, которые способствуют гибкой адаптации к изменяющимся условиям, изобретательному использованию актуальных условий для личностного роста и способности упорно достигать долгосрочных целей в сочетании со способностями к изменению и поднастройке. То есть резильентность подразумевает пластичность.
Пластичность основывается на нейробиологических сетях, которые обеспечивают социальную адаптацию, физическую выносливость и устойчивость за счет гибкого приспосабливания к различным условиям128,129. По-видимому, наиболее пластичным формирование связей является у млекопитающих. Большая нейропластичность привела к развитию живорождения (внутренней беременности) и физиологической реорганизации системы «мать–ребенок», что сделало возможным созревание плода в материнском теле130. Также огромная нейропластичность необходима для того, чтобы сделать новорожденного наиболее значимым объектом для матери по сравнению со всем остальным131.
Как уже было отмечено, окситоциновая система играет ключевую роль в нейропластичности, имеющей решающее значение для формирования привязанности, а послеродовой период соответствует наибольшей пластичности мозга взрослого человека132.
Компонент резильентности «пластичность» включает в себя две особенности: а) резильентность является интегративной и регулирующей; б) резильентность связана со временем.
Резильентность является интегративной и регулирующей
Регуляция способствует гибкой интеграции компонентов системы в функциональное целое, формируя «я», индивидуальность, свободу действий и благополучие посредством формирования новых, специфичных для человека, специфичных для диады и специфичных для культуры конфигураций. Многие исследования проблем развития были направлены на понятие «регуляции», причем некоторые полагают, что это наиболее важная концепция в понимании нарушений развития133,134.
Во многих областях «регуляция» имеет системный уровень. Он определяет, как различные компоненты системы динамически объединяются в функциональное целое, как высшие и низшие элементы иерархически организуются во времени и как компоненты внутри системы интегрируются в режиме реального времени с компонентами ближайшего окружения123,135-137. Концептуальные модели предполагают, что созревание регуляторных процессов происходит от биологических к эмоциональным, затем к управлению вниманием и, в завершение, к саморегуляции138, а совместная регуляция «родитель–ребенок» (синхронность) обеспечивает созревание регуляторных навыков более высокого порядка, таких как управление вниманием и самоконтроль139-141.
Резильентность связана со временем
Резильентность ограничена во времени, имеет процессуальный характер и развивается от простого к сложному, от биологического к психическому. «Временной» компонент резильентности имеет решающее значение не только в процессе эволюции (в филогенезе) и от младенчества до взрослой жизни (в онтогенезе), но и на уровне конкретного социального опыта.
Социальные контакты всегда разворачиваются в рамках временного промежутка, когда два или более участников создают новый «танец» из совпадений и несовпадений, которые определяют поведение, физиологические реакции и психическое состояние участников. Временный характер этих встреч позволяет формировать новые формы из существующих единиц. Следовательно, время является неотъемлемым компонентом резильентности (способности к перенастройке), и это отражается в «синхронности», конструкте, связанной со временем.
Резильентность является социальной
Социальность лежит в основе выживания и адаптации, а виды, которые лучше используют социальные механизмы для координированных действий, имеют значительное преимущество в выживании. Это элегантно описано энтомологом E. Wilson142 в работе «Социальное завоевание земли» («The social conquest of earth»), где он утверждает, что люди достигли превосходства среди позвоночных, а муравьи – среди беспозвоночных, с точки зрения численности населения, распространения по всей земле и выносливости, благодаря их эусоциальности (гиперсоциальности), которая включает в себя способность к совместным действиям членов группы и передачу социальной организации от поколения к поколению.
Примитивные механизмы синхронности обнаруживаются у муравьев, рыб и птиц, при этом координация биологических и поведенческих процессов обеспечивается вазотоцином, родительской молекулой окситоцина и вазопрессина млекопитающих32,62,143. Таким образом, биоповеденческая синхронность у человека опирается на долгую историю социальных механизмов, которые способствовали резильентности путем координации действий. В соответствии с поведенческим принципом аффилиативной нейробиологии эти механизмы прошли отбор с акцентом на поведение: социальное поведение в группе у видов, не относящих к млекопитающим, и аффилиативные связи у млекопитающих. Однако примечательно, что, хотя одиночество опасно для благополучия любого живого организма144, «социальный» компонент резильентности сильно варьируется, и наблюдается широкая изменчивость по всему животному царству, что сочетается с большой изменчивостью плотности и локализации рецепторов окситоцина145,146.
Социальная моногамия
Социальная моногамия знаменует собой первое расширение связи «мать–ребенок» до других привязанностей в семье, в частности, формирования пары и отцовства. Исследования социальной моногамии проводились у нескольких видов приматов (тамарины, мартышки и лемуры) и пяти видов грызунов, происходящих из одной линии (степные полевки, китайские полевки, калифорнийские мыши, карликовые хомяки Кэмпбелла и монгольские песчанки)148.
Моногамия обеспечивает основу для отцовства. Целенаправленное родительское поведение встречается в основном у социально-моногамных видов149. У них отцовство развивается параллельно с материнским уходом, а родители координируют свою заботу по отношению друг к другу150. Участие отца улучшает резильентность молодых млекопитающих, повышает выживаемость потомства, увеличивает размер помета и темпы роста151-156.
Хотя конкретные экологические причины, которые привели к формированию модели совместного воспитания с участием двух родителей у людей, неизвестны, участие отца в воспитании стабилизировало моногамные партнерские системы. После того, как социальная моногамия устанавливается у вида, она способствует появлению сложных социальных форм поведения, которые улучшают резильентность154,157.
Как отцовство, так и парные связи представляют собой расширение связи «мать–младенец», перепрофилирование одних и тех же нейронных сетей и молекулярных процессов и первое проявление последовательной и многообразной нейробиологии аффилиации. Моногамия также требует координации трех внутрисемейных привязанностей (материнство, отцовство и парная связь) в семейную единицу, и такая координация проложила эволюционный путь к человеческой семье и в конечном итоге сложной социально-культурной организации, что обеспечило наилучшую резильентность людей в животном мире.
У людей участие отца в воспитании обеспечивает значительную резильентность. На протяжении всей истории человечества отцы были основным источником косвенной помощи, контролировали материальные ресурсы, физические условия и социальный статус, в которых развивались дети158,159. Исторические отчеты указывают на тесную связь между наличием родителей и детской смертностью в доиндустриальных США и Европе160, а антропологические исследования показывают, что мужчины с большим количеством земли или более высоким социальным статусом демонстрируют больший репродуктивный успех161,162.
В современных обществах более активное участие отца в воспитании повышает резильентность детей, улучшает их психическое здоровье, способствует более высоким академическим и профессиональным достижениям, а также улучшает способность к саморегуляции163,164. Дети, чьи отцы участвовали в воспитании, менее агрессивны и склонны разрешать конфликты, сохраняя уважение, и вести диалог165, а эпидемиологические исследования показывают, что дети, росшие без отца, более склонны к агрессии, нарушению закона и проблемному поведению166,167.
Сложные социальные организации
Хотя социальная моногамия представляет собой первое расширение связи между матерью и ребенком до семейной единицы, считается, что сложная иерархическая социальная организация развивается только у гомининов и растет параллельно с увеличением размера мозга168. Недавние исследования западных горилл обнаружили иерархическую социальную модульность, определяющую не только сложное аффилиативное поведение в рамках обширных родственных групп, но также взаимное сотрудничество между неродственными группами в рамках сезонных коалиций, имеющих общую цель, аналогичных структуре маленькой деревни у людей169.
Такие организации обеспечивают совместный сбор широко рассредоточенных продуктов питания и защиту от хищников, повышая резильентность благодаря совместным действиям вне семьи. Среди приматов, живущих в группах, таких как шимпанзе, наблюдается постконфликтное примирение, которое позволяет членам группы полюбовно разрешать конфликты и поддерживать социальные связи, и эти действия, следующие после акта агрессии, приводят к увеличению окситоцина в моче170.
Исследование на мартышках показало, что чем крепче связь между аффилиативной парой (того же или противоположного пола), измеряемой с точки зрения длительности отношений, времени, проведенного вместе, и проявлений аффилиативного поведения, тем выше эндокринная синхронность колебаний окситоцина в моче171, что указывает на наличие у приматов биоповеденческой синхронности, предшествующей таковой у людей.
Биоповеденческая синхронность – механизм, специфичный для человека
Основываясь на этих социальных механизмах, которые обеспечивают совладание со стрессом, групповую сплоченность и сенсорно-моторную координацию, биоповеденческая синхронность представляет собой специфичный для человека механизм, посредством которого два человека могут взаимно влиять на физиологию друг друга без физического контакта, посредством координации лицевых аффективных сигналов, которой нет у других приматов и грызунов29.
У человека синхронность развивается в течение всей жизни и постепенно усложняется, включает совместное построение общего нарратива, способность принимать различные точки зрения и способность сопереживать боли, действиям, эмоциям и психическому состоянию другого человека. Развитие синхронности начинается с распознавания матерью биологических ритмов младенца в своей утробе и заканчивается построением близких отношений между взрослыми, наполненных взаимной заботой.
Резильентность включает в себя осмысленность
В то время как первые два принципа резильентности основаны на механизмах, общих для всех видов, смыслообразущий компонент является специфичным для человека. Для формирования концепции резильентности у человека мы должны объединить общие для всех видов механизмы выносливости, многообразия, адаптации и совладания со стрессом со способностью человека придавать значение трудностям, невзгодам и травмам.
Люди способны придавать смысл травме, используя коллективные культурные или религиозные мифы, или в других случаях, наделяя персональными смыслами особые действия, обычно такие, которые ведут к укреплению аффилиативных связей, или акты альтруизма, выходящие за пределы личности.
Во многих исследованиях подчеркивается роль духовности в способности восстанавливаться после перенесенных невзгод или использовать травму для роста172-174. Исследования также указывают на важность щедрости для резильентности175 и на утешительную функцию религиозных сообществ, которые придают коллективный смысл, а также оказывают поддержку176,177. W. James в работе «Воля к вере»178 («The will to believe») рассматривал веру как преднамеренный выбор, который дает опору и позволяет человеку сформировать личностно-смысловой взгляд на реальность, придавать значение травме и трудностям. Его знаменитая метафора превращения дискретного опыта в осмысленное целое – «живые электрические провода», которые светятся и сияют, в отличие от «мертвых проводов», которые остаются расплывчатыми и тусклыми, – элегантно описывает эту функцию веры, способствующую резильентности.
Создание смысла вводит дименсию «будущее» в концепцию резильентности, расширяя временной горизонт за пределы «помнящего присутствия»179 других приматов. Это подчеркивает целенаправленную функцию культурных мифов, которые создают люди и которые выходят за рамки жизни человека и подпитывают внутренние резервы резильентности перед лицом трудностей.
Обретение смыслов, выходящих за рамки индивидуальной жизни, не только является ключевой особенностью резильентности, но и находит объяснение в двух нейробиологический системах аффилиации. Carter76 предположил, что окситоциновая система обеспечивает нейробиологический субстрат для духовности благодаря своей роли в поддержании любви, заботы, эмпатии и морального подъема, в частности потому что позволяет млекопитающим испытывать «состояние бдительности без страха», что значит «быть полностью в настоящем моменте без ощущения потенциальной опасности». Точно так же нейронные структуры, связанные с «аффилиативным мозгом» и формирующиеся в ранние сензитивные периоды, позволяют людям распространять любовь на незнакомцев, социальные группы и абстрактные идеи, проявлять великодушие за пределами непосредственных связей индивида.
Тем не менее сильные культурные мифы разных поколений, системы смыслов и религиозные верования способны нивелировать первый принцип резильентности – гибкость – путем ужесточения обычаев, обязательств, требуя подчинения, ужесточая контроль. Такие группы часто функционируют благодаря тесной внутренней сплоченности, достигаемой за счет усиления нейронной и поведенческой синхронности между ее членами до гиперсоциального уровня перед лицом реальной или мнимой опасности. Например, на протяжении всей истории человечества солдаты проходят интенсивную подготовку к скоординированным действиям, и эта моторная синхронность позволяет устранить когнитивную эмпатию во время битвы, чтобы сражаться и уничтожать тех, кто к группе не относится. Социальный принцип резильентности становится более значимым для группы и неприменим к тем, кто вне ее.
Примечательно, что и окситоцин, и нейрональная синхронность задействованы в таком разделении «относящийся к группе / не относящийся к группе», построенном на древних механизмах, позволяющих немедленно отличить друга от врага, чтобы защитить близких. К примеру, мы изучали нейронную реакцию израильской и палестинской молодежи с использованием магнитоэнцефалографии (МЭГ), когда они наблюдали за тем, как переносили боль представители их группы и те, кто к ней не относился. В течение первых 500 мс, представляющих автоматический ответ мозга на чужую боль, молодые люди реагировали на боль и «своих», и «чужих». Однако спустя эти полсекунды сопереживания высшие процессы блокировали естественную эмпатическую реакцию мозга на боль представителя другой группы, давая проявиться только реакции на боль представителя своей группы180.
Два процесса помогали отключить эволюционно-древнюю эмпатическую реакцию на конспецифическое состояние при дистрессе: повышение уровня окситоцина и усиление нейронной синхронизации между членами группы. Таким образом, окситоцин и нейронная синхронизация определялись системой смыслов, которая подавляла эмпатию к тому, кто не был членом группы.
Исследование роли окситоцина в отвержении «чужих»181, ставит вопрос, как интегрировать роль систем смыслов, которые, способны, с одной стороны, улучшать резильентность путем создания сообществ и придания травме межпоколенческого значения, а с другой – вызывать агрессию и предубеждение к тем, кто не относится к группе. Возможно, одно из решений заключается в том, чтобы направлять постоянные усилия на то, чтобы придать древним системам смыслов гибкость и человечность, чтобы старые ритуалы не становились ригидными и распространялись на всех людей.
СИНХРОННОСТЬ ОТ МЛАДЕНЧЕСТВА
К ЗРЕЛОСТИ: РАЗВИТИЕ РЕЗИЛЬЕНТНОСТИ
Созревание синхронности происходит не только в ходе эволюции животных, но и на протяжении всей человеческой жизни. Первоначально синхронность проявляется главным образом в отношениях матери и ребенка, первичной связи у млекопитающих, а затем распространяется на другие социальные связи, включая отца, наставников, близких друзей и романтических партнеров, все человечество, проявляется в чувстве синхронности с природой, искусством и священным опытом.
Эти представления предоставляют биологические и научные доказательства концепции Winnicott, изложенной в «Игре и реальности»182 («Playing and Reality»), о непрестанном присутствии матери в качестве основы для формирования символов, игры, творчества и духовного опыта. Синхронность усложняется по мере разнообразия репертуара, перехода к символическому уровню и зависит от степени взаимности в детстве и подростковом возрасте, настраивает социальный мозг по мере получения нового опыта на понимание других, проявление сочувствия к чужим страданиям и участие в отношениях183. Корни синхронности происходят из эволюционно древних паттернов, в биологических ритмах плода эти переживания проявляются на физическом плане, что позволяет резонировать всей истории отношений в моменте встречи человека с человеком.
В то время как философские взгляды на «объединение» предполагают, что «я» строится из микроидентификаций, которые раскрываются во время конкретного повседневного опыта, синхронность добавляет элемент, который «я» берет из конкретных паттернов с участием значимого человека. Наша модель рассматривает подробнее развитие этого феномена как в ходе эволюции, так и на протяжении жизни человека, и показывает его вклад в формирование устойчивости к неблагоприятным условиям.
Синхронность между матерью и младенцем начинается, когда мать распознает первые биологические ритмы младенца в утробе, такие как сердечные ритмы и циклы сна и бодрствования, которые посылают сигналы к плаценте и материнскому мозгу184-186. После рождения матери вовлекают эти знакомые ритмы в диадический обмен.
Исследования 1970-х годов показали, как взаимодействие между матерью и младенцем «лицом к лицу» строится на модели «действие–пауза» повторяющихся биологических актов, таких как сосание или плач187,188. От простого участия в этих биологических ритмах синхронность постепенно развивается в эмпатические отношения между взрослыми, в рамках которых возможен диалог.
Мы изучали проявления синхронности между матерью и ребенком от рождения до 25 лет и наблюдали, как при взаимодействии в течение четверти века сохранялись одни и те же невербальные ритмические паттерны, колебания возбуждения и положительные пики. Например, в некоторых диадах уровень возбуждения постоянно колебался между низким и средним, в то время как в других взаимодействие происходило в виде быстрых пиков положительного возбуждения. Такая стабильность создает базовый порядок, пока более сложные и творческие паттерны встраиваются в диалог и образуют знакомое и единое событие.
Помимо обеспечения «ритма безопасности» для улучшения резильентности особенно важны две дополнительные функции синхронности. Во-первых, микроструктура синхронного опыта – это постоянное чередование разрыва контакта и его восстановления. Согласно Tronick189 матери синхронизируются с младенцем только около 30% времени; таким образом, диады проводят больше времени в неверно скоординированных состояниях, чередующихся с драгоценными моментами синхронности. Сторонники психоанализа и теории развития189,190 подчеркивают важность таких циклов соответствия и несоответствия для обучения младенцев тому, как переживать моменты отсутствия сонастройки и как устранять недопонимание, присущее человеческому диалогу.
Описаны два типа отклонений от паттерна «длительное несоответствие–короткое соответствие». Первое – гиперсинхрония, обнаруживается у тревожных матерей и выражается в увеличении числа эпизодов соответствия (более 45% времени); второе – абстиненция, наблюдаемая у депрессивных матерей, связана с почти полным отсутствием сонастройки. Оба приводят к трудностям саморегуляции у младенцев191,192. Таким образом, синхронность, создает серию микрособытий, состоящих из постоянного чередования разрыва контакта и его восстановления, и в безопасных условиях учит детей переносить фрустрацию. В возрасте около 9 месяцев у младенцев появляется способность самостоятельно восстанавливать контакт193, что готовит их к построению равных отношений с друзьями и партнерами.
Второй функцией синхронности, способствующей резильентности, является ее роль в качестве первого контекста для развития прогностических способностей мозга. Недавние модели «прогнозирующего кодирования» рассматривают мозг как вычислительное устройство, главная роль которого заключается в повышении адаптации путем минимизации энтропии и повышения уверенности. Нейронные волны играют важную роль в кодировании с построением прогноза: альфа-волны участвуют в построении прогноза, бета-волны – в оценке его точности, а гамма-волны – в оценке ошибки прогноза, постоянном точечном соотнесении прогноза с поступающей информацией196,197. Опыт синхронности обеспечивает шаблон для полиритмической когерентности, который создает мультисенсорный образ тела в мире197 и включает в себя интеграцию альфа-, бета- и гамма-ритмов при формировании социальных прогнозов в реальном времени112.
Используя экологическую парадигму и методы гиперсканирования, мы обнаружили связь гамма-ритмов между мозгом матери и ребенка118, романтических партнеров198 в моменты поведенческой синхронности. В исследованиях как у животных199,200, так и у людей201,202 гамма-ритмы показывают уровень зрелости мозга, что указывает на роль синхронности в тонкой настройке этой зрелости. Гамма-ритмы и ошибки прогноза в висцеромоторной коре и областях, ответственных за мотивацию, усиливают чувства, но из-за неоднородности этих областей различия между «я» и другими стирается203. Таким образом, изучение синхронности может помочь по-новому взглянуть на социальное созревание мозга в реальных условиях.
Стадии развития синхронности
По результатам многочисленных продольных и поперечных исследований мы выявили пять различных этапов развития синхронности от периода беременности до юности и показали наличие индивидуальной стабильности между этими стадиями и ее чувствительность к определенным неблагоприятным условиям.
Подготовка к синхронности связана с постепенным знакомством матери с биологическими ритмами плода: циклами сна–бодрствования, консолидирующимися примерно на 31–32-й неделе беременности204,205, за которым следует организация сердечных ритмов примерно на 33-й неделе беременности206,207. Эти циклы согласуются с реакцией плаценты208, и хорошая организация этих биологических ритмов прогнозирует лучшую синхронность между матерью и младенцем в 3 месяца121.
Период новорожденности: поведение матери в послеродовом периоде
Сразу после рождения и в течение первых 6 недель жизни человеческие матери, как матери всех млекопитающих, проявляют видоспецифичный репертуар материнского поведения, который у людей включает в себя рассматривание лица и тела младенца, выражение положительных эмоций, высокие вокализации «материнского языка» и нежные прикосновения. Однако в отличие от других млекопитающих у людей матери координируют свое поведение с краткими моментами настороженности новорожденного. Таким образом, в норме при рождении у младенца происходит координация между его внутренним состоянием и реакцией социального мира.
Материнское послеродовое поведение в неонатальном периоде обеспечивает основу для развития символической компетентности209, лучшего когнитивного развития и меньшего количества экстернализированных и интернализированных симптомов в раннем детстве210 и коррелирует с уровнем окситоцина у родителей211.
Младенчество: аффективная синхронность
С третьего месяца жизни мать и младенец начинают участвовать в интерактивном «танце», создают специфичный для диады ритмичный диалог, используя взгляды, выражение эмоций, ко-вокализации и прикосновения. Этот невербальный опыт играет ключевую роль в социальном, эмоциональном, когнитивном развитии ребенка114. Матери и отцы участвуют в специфичных для родителей формах синхронности, более ритмичных у матерей и объект-ориентированных у отцов212.
Аффективная синхронность между родителями и детьми связана со множеством гормонов, питающих привязанность, таких как окситоцин, вазопрессин, бета-эндорфин, пролактин, кортизол и слюнная альфа-амилаза, а также с иммунными биомаркерами, включая IgA и IL-6 в слюне213. Помимо этого, она связана с активацией аффилиативного мозга у матерей214 и отцов89. Невербальная синхронность также обнаруживается при взаимодействии триады «мать–отец–ребенок»215, что создает основу для социального участия детей в отношениях внутри группы и культуры.
Преддошкольники/дошкольники: символические игры и совместное конструирование воображаемых нарративов
На втором и третьем годах жизни дети начинают участвовать в символической игре и наполнять объекты символическим значением и «похожими на рассказ» символическими последовательностями. Синхронность с матерью и отцом в младенчестве216 не только определяет уровень сложности символов, также организацию символических игр переопределяет временной контур ритмического обмена младенца с матерью и отцом – коротких, случайных и многочисленных интеракций с отцом и более редких и длительно выстраиваемых – с матерью126.
В дошкольные годы дети начинают совместно строить диалог, используя будущее и прошлое время, воображаемые сценарии и альтернативную реальность, в которую они могут погрузиться. Эти игровые творческие способности опираются на невербальную синхронность первых месяцев жизни210 и превращают синхронизированный диалог в социальное событие, включающее в себя творчество, использование языка и деятельность формирующегося сознания, которые выражают внутреннюю реальность. Взаимодействие дошкольников с матерью и отцом предопределяет умственные способности детей и развитие нравственной позиции в детстве и подростковом возрасте114.
На этом этапе у детей появляются «лучшие друзья», и они вступают в социальные институты. Опыт аффективной синхронности формирует социальные компетенции ребенка со сверстниками в зависимости от культурных традиций217. Уровень родительского окситоцина, гены OXTR и ранняя синхронность позволяют прогнозировать синхронность детей с их первым лучшим другом72.
Позднее детство / подростковый возраст: эмпатический диалог
Начиная примерно с 9–10 лет и вплоть до подросткового возраста, дети заметно сокращают количество «игровых» взаимодействий со своими родителями, и диалог приобретает словесную форму – вид взаимодействия, который предполагает разрешение конфликтов, обмен информацией и, в норме, обсуждение ребенка с родителями пережитого опыта, идей, чувств, обмен мнениями и планами на будущее.
Синхронизированный диалог на этом этапе запускает у ребенка способность к поведенческой, эмоциональной и когнитивной эмпатии, умение планировать заранее, разрабатывать, сотрудничать, проявлять мотивацию и понимать точку зрения другого человека. Такие социальные способности, особенно на этой стадии, когда фокус привязанности переносится с родителей на друзей, имеют решающее значение для благополучия детей и связаны с устойчивостью перед лицом трудностей и созреванием социального мозга113,218-220.
Взрослая жизнь: взаимность, близость и взаимопонимание
Если связь между матерью и ребенком была «достаточно хорошей» и синхронность развивалась в соответствии с возрастными нормами, создавая пространство как для согласованного контакта, так и для фрустрации, мать и выросший ребенок могут встретиться друг с другом как двое взрослых, которые все еще сохраняют свои роли, но могут вести диалог с уважением к зрелости друг друга, который будет отражать их отношения в целом. Он опирается на ранние привычные ритмы и перекликается со всеми этапами развития, но этот диалог, который является взаимным и уважительным, интимным и автономным, знакомым и безопасным, все же отличается от диалога с партнером.
Такая надежная синхронность позволяет людям вступать в другие взаимные и доверительные отношения и перекидывает мост к следующему поколению, которое может превзойти родителя благодаря способности повзрослевшего ребенка пробуждать в своем собственном мозгу диадические переживания от взаимодействия с родителем в его отсутствие.
В целом синхронность, которая постепенно обогащает социальный репертуар младенца, обеспечивает созревание более сложных умственных способностей, открывает путь изменчивости при сохранении порядка, разнообразию при сохранении стабильности. Синхронность касается той области жизни, где биологическое объединяется с социальным, чтобы придать смысл, сформировать узы и противостоять трудностям.
ВОСПИТАНИЕ РЕБЕНКА С ХОРОШЕЙ РЕЗИЛЬЕНТНОСТЬЮ: ЛОНГИТУДИНАЛЬНОЕ ИССЛЕДОВАНИЕ ТРЕХ КОГОРТ ВЫСОКОГО РИСКА
Наша модель предполагает, что биологические и социальные положения, заложенные в связи между матерью и ребенком, обеспечивают основу для резильентности на протяжении всей жизни. Однако у многих детей по всему миру эти положения оказываются нарушены. Чтобы улучшить понимание резильентности, мы должны рассмотреть неблагоприятные факторы в отдельности, изучить, что именно нарушается в каждом отдельном случае, и проверить, как эти упущения влияют на результат.
Мы предположили следующее: исследования на людях должны начинаться с рождения или сразу после него; предпочтителен лонгитудинальный дизайн; изучение «недостающего компонента» должно строиться с учетом данных конкретных исследований на животных моделях, выявивших эти положения, путем оценки их последствий для детского мозга и поведения30.
Существуют три основных источника нарушений детско-материнских отношений: связанные с матерью, ребенком и условиями среды. Каждый из них затрагивает миллионы детей во всем мире.
Послеродовая депрессия отмечается у 15–18% рожениц в индустриальных обществах и достигает 30% в развивающихся странах221. Мы предположили, что работа Meany222 о долгосрочном влиянии недостаточного ухода (в виде вылизывания и груминга) у крыс на выработку окситоцина и реакцию на стресс у детенышей может дать представление о долгосрочных последствиях материнской депрессии.
Преждевременные роды происходят в 10,5% живорождений в индустриальных обществах223, и хорошо известен их негативный эффект, в частности связанный с разлукой с матерью в период выхаживания и влиянием на системы жизнеобеспечения, что согласуется с моделью «материнской близости» Hofer27.
Стресс на ранних этапах жизни имеет долгосрочные негативные последствия для развития. Каждый пятый ребенок в мире растет в условиях хаоса, иммиграции, отсутствия продовольственной безопасности или жилья, племенной или этнической войны, бедности и насилия. Животная модель, которую можно считать аналогичной, предполагает «варьирование доступности кормовой базы»224,225 у индийских макак, в которой матерям пища то доступна, то недоступна или труднодоступна, и таким образом для детеныша непредсказуемым образом меняются периоды, когда мать заботиться о нем, и периоды, когда забота сводится к минимуму. Установлено, что такие условия оказывают худшее влияние на потомство – с точки зрения развития мозга, реакции на стресс и поведения – по сравнению с постоянными хорошими или плохими условиями жизни, что позволяет предположить, что непостоянство условий в раннем периоде жизни является наиболее вредным для резильентности детей.
Чтобы понять резильентность с точки зрения нейробиологии развития, мы исследовали три когорты матерей и младенцев в период от рождения (или младенчества) до подросткового возраста / ранней молодости, уделяя особое внимание тому, как нейробиологические компоненты аффилиации отличались у детей в зависимости от жизненного пути. В каждой группе имелось одно из вышеупомянутых нарушений детско-материнской связи, и гипотезы основывались на аналогичных животных моделях.
В когорту послеродовой депрессии были включены дети, матери которых страдали хронической депрессией в течение первых лет жизни ребенка. Когорта, живущих в условиях войны, включала матерей и детей, живущих в зоне непрерывных боевых действий, а группа недоношенных включала неврологически благополучных недоношенных детей, у половины из которых в неонатальном периоде был обеспечен контакт «кожа к коже» («метод Кенгуру»).
В течение всего детства проводилась повторная оценка показателей синхронности, навыков саморегуляции, уровня окситоцина, гормонов стресса, уровня психического развития и в конечной точке – нейровизуализационное исследование социального мозга.
Послеродовая депрессия
В нашу когорту были включены только матери, физически здоровые, имеющие партнера, старше 21 года, чей доход был выше черты бедности, чтобы отделить проявления собственно депрессии от часто сопутствующих негативных факторов (матери-одиночки, матери-подростки, бедность). Оценка депрессии проводилась неоднократно в течение первого года и снова через 6 и 10 лет. Мы сформировали две когорты: дети матерей, которые страдали хронической депрессией от рождения до достижения ребенком 6 лет, и здоровый контроль.
Послеродовая депрессия у матери увеличивает риск психических заболеваний
Наличие депрессии у матери заметно повышает вероятность психических расстройств у детей, даже в семьях с низким риском. В возрасте шести лет у 60% детей матерей, у которых было диагностировано большое депрессивное расстройство как в 9 месяцев, так и в 6 лет, и которые сообщали о том, что в целом депрессия наблюдалась в первые годы жизни ребенка, был установлен психиатрический диагноз Оси I (по сравнению с 15% в контрольной группе), причем наиболее распространенными расстройствами были тревожное расстройство и расстройства поведения226. В 10 лет и в подростковом возрасте более, чем у 50% этих детей все еще сохранялся психиатрический диагноз, даже когда у матери отмечалась ремиссия, что указывает на долгосрочный эффект такого раннего воздействия. Выраженность экстернализированых и интернализированных симптомов была больше у детей матерей, страдающих депрессией227.
Синхронность способствует резильентости
Матери, страдающие депрессией, не могли обеспечить ребенку соответствующий уход, необходимый для развития. В 9 месяцев микроанализ невербального поведения показал у матерей с депрессией минимальные показатели зрительного контакта, положительного аффекта и нежных прикосновений и минимальную синхронизацию со своим ребенком191. Поскольку синхронность развивается с течением времени, матери, страдающие депрессией, не могли развить более зрелые формы взаимного диалога.
Синхронность была индивидуально стабильной от рождения до подросткового возраста, и более низкая синхронность у детей матерей с депрессией сочеталась с развитием психических заболеваний и социальной отгороженностью у детей. В 6 лет дети матерей, страдающих депрессией, демонстрировали недостаточную поведенческую эмпатию228. В 10 лет они демонстрировали более низкие исполнительские функции и недостаточное распознавание эмоций. Более низкая синхронность предопределяла снижение показателей социально-эмоционального интеллекта.
В то же время синхронность функционировала как компонент резильентности. Если дети матерей с депрессией все же получали больше синхронности (от своих отцов, из-за большей активности системы окситоцина или из-за врожденной общительности ребенка), это служило защитным фактором. Способность детей функционировать более адекватно в социальном мире, создавать дружеские отношения и участвовать во взаимодействиях со сверстниками, опосредованных синхронностью, заметно снижает риск психических расстройств, уменьшает влияние материнской депрессии в ранние годы ребенка на его исполнительские способности и распознавание эмоций. Этот эффект был особенно заметен в позднем детстве, в период, когда отношения между сверстниками начинают оказывать большее влияние на жизнь детей. Это подтверждает наше утверждение, что компоненты резильентности функционируют по-разному на разных этапах и что аспекту развития должно уделяться основное внимание в концепции резильентности.
Измененная реакция на стресс опосредуется негативным воспитанием
Эффекты материнской депрессии на стрессовую реакцию у детей были разнообразными, меняясь в зависимости от стадии развития, используемых методик оценки и показателей резильентности. В 9 месяцев у младенцев матерей, страдающих депрессией, наблюдалась повышенная реактивность кортизола при воздействии социального стрессора и замедление его нормализации229. В 6 лет сохранялось влияние материнской депрессии на вариабельность кортизола, но только среди детей, которые воспитывались в напряженных, угрожающих и негативных условиях228. Эти результаты подчеркивают важность компонента пластичности/гибкости для стрессоустойчивости. В 10 лет только дети матерей с депрессией, которые воспитывались в негативных условиях, имели более высокий уровень кортизола, такая чрезмерная активация оси гипоталамус-гипофиз-надпочечники (ГГН) опосредовала влияние депрессии на развитие психопатологии221.
Мы также измеряли IgA в слюне, биомаркер иммунной системы, и обнаружили более высокие показатели у детей матерей, страдающих депрессией, в позднем детстве, что указывает на больший стресс. Однако это было выявлено только среди детей, чья синхронность с матерью была минимальна, что свидетельствует о влиянии синхронности на стрессоустойчивость и иммунную систему.
Отцы повышают резильентность
В условиях минимальной синхронности, обеспечиваемой матерью в депрессии, хорошо синхронизированные отношения с отцом выполняли важную функцию в формировании резильентности. Если отцы относились к детям внимательно и отзывчиво, риск развития психических расстройств заметно снижался230. Похоже, что одним из механизмов, с помощью которого заботливое отцовство улучшает резильентность, является изменение семейной атмосферы, что делает семейные взаимодействия более согласованными, гармоничными и вовлеченными, даже когда матери находятся в депрессии231. Эти результаты согласуются с механизмом «социальной моногамии», описанным выше, и предполагают, что связь с матерью открывает ребенка другим аффилиативным связям в семье и обеспечивает резильентность.
В другом исследовании мы наблюдали за родителями и их первенцами в израильском и палестинском обществе от младенчества до дошкольного возраста. Мы обнаружили, что материнская депрессия оказывает менее токсичное влияние на психику и символические компетенции ребенка в палестинском обществе, что было связано с условиями жизни в расширенной семье в этой культуре, которая предоставляла детям широкие возможности для синхронного взаимодействия с другими взрослыми родственниками232.
Окситоцин способствует резильентности
Через 6 и 10 лет у матерей с депрессией и их детей был снижен уровень окситоцина как в слюне226, так и в моче233. Как у матерей, так и у детей распространенность генотипа GG по гену OXTR (rs2254298) была выше, что связано с большей уязвимостью для психических расстройств234. Если у матери имелся аллель А в гене OXTR, вероятность установления диагноза по оси I у ребенка в 6 лет снижалась вдвое226. В 10 лет, если уровень окситоцина в слюне у детей был высоким, влияние материнской депрессии на экстернализированные и интернализированные симптомы ребенка было меньше227.
Аффилиативный мозг подростков
В раннем подростковом возрасте мы оценивали нервную эмпатическую реакцию детей на боль других и привязанность с помощью МЭГ. У детей матерей, страдающих депрессией, мы обнаружили нарушения нервного эмпатического ответа в верхней височной борозде, которая является центром социального мозга, проявившиеся в снижении альфа-активации и более быстром прерывании нервного ответа примерно через 900–1100 мс после стимула. Такое прерывание ответа сочеталось с интрузивным и негативным воспитанием детей и недостаточной синхронностью, которую эти подростки испытывали в младенчестве, что усугубляло пагубные последствия влияния материнской депрессии для развития мозга219.
Для нейробиологической оценки привязанности мы использовали типичный тест – сравнение реакции детей на демонстрацию их взаимодействия с матерью на более ранней стадии и взаимодействия с незнакомцами. Типичная нейронная активация на сигналы привязанности включала мультиритмический альфа-, бета- и гамма-ответ, проявившийся в альфа-супрессии в задней области, и бета- и гамма-активации в обширном кластере справа, включавшем верхнюю височную борозду, веретенообразную извилину и островок. Тем не менее только те дети матерей с депрессией, у кого развилось аффективное расстройство, демонстрировали аберрантный ответ, включающий сниженную реакцию на социальные сигналы и ослабление дифференциации между стимулами привязанности и непривязанности. Эти нарушения предопределялись снижением активности системы окситоцина и снижением синхронности между матерью и ребенком в детстве.
Хотя эти данные указывают на риск нарушений последующих привязанностей у детей матерей, страдающих депрессией, они также показывают, что некоторые дети, растущие в условиях хронической материнской депрессии, более устойчивы, и что нейробиологические компоненты аффилиации являются маркерами резильентности.
Стресс и психотравма в раннем детстве
Когорта переживших стресс и психотравму в раннем возрасте включала детей и их матерей, живущих в зоне непрерывных боевых действий, которые подвергались повторным непредсказуемым ракетным обстрелам в течение почти 20 лет. Обследование детей проводилось в младенчестве, среднем детстве (5–7 лет) и позднем детстве (10 лет), в раннем подростковом возрасте выполнялось нейровизуализационное исследование социального мозга.
Коморбидные психические расстройства у детей с хронической травмой, полученной в раннем возрасте
У детей, растущих в таких травмирующих и непредсказуемых условиях, выявлено увеличение распространенности психических расстройств по оси I в 3–4 раза и заметное усиление интернализированных и экстернализированных симптомов. В сравнении с когортой детей, чьи матери находились в депрессии, особенность этой когорты заключалась в том, что две трети детей имели более одного диагноза, причем у некоторых было три или даже четыре психических расстройства, что свидетельствует о том, что травма ведет к множественным дисфункциям по всему психопатологическому спектру235.
В ходе анализа резильентности в течение первого десятилетия жизни у детей, проживающих в травматичных условиях, мы обнаружили, что матери детей, у которых не было выявлено психических расстройств или отмечалось выздоровление, демонстрировали меньшую выраженность симптомов и проявляли большую синхронность. Такие дети проявляли большую социальную компетентность в позднем детстве (10 лет)236.
Окситоцин как буфер стресса
В исследуемой группе активность окситоцина была связана с резильентностью при травме. Большая активность гена рецептора окситоцина у детей, матерей и отцов отличала детей, у которых развился хроническое посттравматическое стрессовое расстройство (ПТСР), от детей, у которых диагноз был снят в среднем детстве235.
В возрасте 10 лет, в отличие от детей матерей, страдающих депрессией, групповых различий по уровню окситоцина не было выявлено, что свидетельствует о том, что не все дети, растущие в зоне военных действий, демонстрируют фундаментальные биологические нарушения аффилиации и что некоторые матери способны, прилагая значительные усилия, смягчить опасные последствия войны для своего ребенка. Однако уровень окситоцина у матерей, живущих в условиях войны, был ниже, что свидетельствовало об огромном напряжении при воспитании ребенка в травматичных и непредсказуемых условиях. Множество таких изменений затрагивают эндокринную и нервную систему матери.
Выявлена эндокринная синхронность между матерью и ребенком. Когда уровень материнского окситоцина был низким и снижался уровень синхронности, у детей проявлялось значительно больше симптомов. Но этого не наблюдалось в случаях, когда у матери сохранялся высокий уровень окситоцина и стиль воспитания был заботливым и неинтрузивным120.
Реакция на стресс
Мы измеряли базальный уровень кортизола и его колебания у матери и ребенка в раннем детстве, в позднем детстве и в раннем подростковом возрасте. Уровень гормонов оценивался в волосах и слюне. В раннем детстве уровень кортизола и альфа-амилазы слюны, маркера симпатической реакции на стресс, различались у детей с ПТСР и без него. У детей без ПТСР уровень гормонов был значительно выше, в то время как у детей с ПТСР отмечались более низкие уровни236. Эти данные свидетельствуют о том, что в условиях хронической травмы в раннем детстве большая активация оси ГГН способствует резильентности, а не увеличивает риски.
Через 10 лет базальные показатели и колебания гормонов оси ГГН были повышены только у детей, проживавших в условиях военных действий, у которых развились психические расстройства, и это были дети матерей с более выраженной активацией оси ГГН и меньшей синхронностью в детско-родительских отношениях73. Мы предполагаем, что «матери стояли между войной и ребенком», и в случаях, когда мать способна совладать с собственным стрессом и защищать ребенка от внешней травмы, она способна сдерживать стрессовую реакцию ребенка.
Тем не менее в раннем подростковом возрасте дети и их матери в целом демонстрировали более высокие и менее вариабельные уровни кортизола, что позволяет предположить, что хроническое воздействие непредсказуемого стресса само по себе является фактором риска, независимо от взаимоотношений. Возможно, такая уязвимость сопровождает ключевые периоды развития, в частности переход к подростковому возрасту237. Показатели иммунных биомаркеров были выше у матерей и детей, проживающих в зоне боевых действий, что подчеркивает значительное истощение иммунной системы в условиях хронического стресса и согласуется с моделями аллостатической нагрузки и реакции на стресс238.
Мозг ребенка и матери
В этой когорте, в отличие от двух других, мы рассматривали мозг матери и ребенка в идентичных условиях, пытаясь оценить, как хронический стресс воздействует на нервные системы обеих сторон диады. Мы обнаружили, что изменения в функционировании мозга были опосредованы историей взаимоотношений, что привело к различному (благополучному и неблагополучному) созреванию социального мозга у детей.
Мы оценивали коннективность и мощность сети пассивного режима работы головного мозга (СПРРГМ, default mode network – DMN), обеспечивающей чувство самосознания, переключения внутреннего и внешнего внимания и автобиографическую память239-241. И у матерей, и у детей были обнаружены и нарушения коннективности СПРРГМ, и ее мощности, что еще раз подчеркивает роль пластичности в формировании резильентности и указывает на недостаточную способность отдельных структур объединяться в единую систему, которая обеспечивает основу самосознания.
У матери обнаруженные нарушения СПРРГМ происходили на уровне альфа-ритма, основного ритма бодрствующего зрелого мозга, тогда как нарушения СПРРГМ у детей происходили в тета-диапазоне, биомаркере развивающегося мозга242. У детей с ПТСР наибольшие нарушения затрагивали тета-соединения. Нарушение паттернов тета-связности было опосредовано материнским интрузивным, провоцирующим тревогу воспитанием и более высоким уровнем кортизола в более позднем детстве, что подчеркивает долгосрочные негативные последствия непредсказуемого воспитания, при котором родитель не управляет своими эмоциями, для ключевой нейронной системы243.
У детей не было обнаружено никаких групповых различий нейронального эмпатического ответа на страдания других. Этот ответ приводил к альфа-активации большого кластера, включавшего дополнительную моторную область, часть нейросети, отвечающей за самосознание, и срединную поясную извилину, узел СПРРГМ. Синхронность, которая была снижена в когорте живущих в условиях боевых действий, опосредовала влияние ранней травмы на нервный эмпатический ответ, и дети, получившие больше синхронности в детстве, демонстрировали более активную реакцию на страдания других244. Нервная эмпатическая реакция матерей также имела нарушения, но они были специфическими для мозга взрослого245.
Недоношенность
Наш проект с использованием «метода Кенгуру» является единственным существующим исследованием, в котором оценены эффекты разлуки с матерью, отсутствие структурированного контакта с ней на созревание функций жизнеобеспечения у младенцев в долгосрочной перспективе. Матери недоношенных детей с низкой массой тела при рождении (<1750 г) были рандомизированы в две группы. В одной применялась экспериментальная методика (контакт «кожа к коже» не менее часа в день не менее 14 дней подряд в течение периода выхаживания), в другой – выхаживание младенца осуществлялось стандартным способом в инкубаторе.
Диады обследовали семь раз в течение первого десятилетия (до вмешательства, при выписке, в возрасте 3, 6, 12 и 24 месяцев, в 5 и 10 лет). По достижении детьми молодого возраста (18–20 лет) мы посещали их на дому, чтобы оценить их отношения с матерями, гормональные показатели и исполнительские функции, а в течение следующего месяца проводили функциональную магнитно-резонансную томографию.
Мы обнаружили, что обеспечение телесного контакта с матерью у людей влияло на те же системы, что и у других млекопитающих. «Метод Кенгуру» улучшал у новорожденных функционирование вегетативной системы, организацию циклов сна–бодрствования, ориентацию и обработку информации. В то же время он обеспечивал потребность в материнской заботе в период новорожденности246.
В соответствии с нашей моделью поэтапного развития регуляторных функций138 эти улучшения в физиологической регуляции и взаимодействии с матерью способствовали улучшению резильентности и влияли на динамику развития. Новорожденные демонстрировали лучшую модуляцию возбуждения при обработке сильных возбуждающих стимулов в 3 месяца, лучшее исследовательское поведение в 6 месяцев и лучшие способности к самоконтролю в 1 и
2 года. Умственные, но не моторные способности были лучше в экспериментальной группе через 6, 12 и 24 месяца247,248. Также улучшалась синхронность между матерью и младенцем, а у матерей увеличивалось количество грудного молока в ответ на увеличение уровня окситоцина249. После использования «метода Кенгуру» более высокая синхронность отмечалась при каждом обследовании в первые годы, и большая взаимность при социальных взаимодействиях была связана с лучшими когнитивными и регуляторными способностями210.
Через 10 лет мы обнаружили, что лучшие регуляторные способности субъектов из экспериментальной группы сохранялись. Мы обнаружили более высокую дыхательную синусовую аритмию и ее большую чувствительность к эмоциональному стрессу, что указывает на более адаптивное функционирование вегетативной нервной системы. Оценка сна выполнялась с помощью актиграфии в течение пяти ночей подряд, и у детей, к которым был применен «метод Кенгуру», организация сна была лучше, периоды бодрствования короче. Кроме того, реакция оси ГГН у детей из экспериментальной группы на социальный стрессор вызывала сниженный кортизольный ответ на стресс и отмечалось более быстрое восстановление250. Что касается когнитивных способностей, к 5 годам больше не было различий общего IQ, но дети, у которых применялся «метод Кенгуру», имели лучшие исполнительские способности, рабочую память и когнитивную гибкость через 5 и 10 лет.
В целом наши результаты позволяют выделить системы, на которые влияют компоненты устойчивости, связанные с материнским телом и адаптивными формами ухода, и подтверждают наличие их связей с совладанием со стрессом, гибкой реакцией на условия окружающей среды, модуляцией возбуждения и внимания, а также способностью участвовать в взаимном диалоге.
В молодом возрасте для оценки эмпатической реакции мозга на эмоции других в группе детей, у которых применялся «метод Кенгуру», и контрольной группе изучали «эмпатическую точность», важную детерминанту эмпатической реакции251,252 и дифференцированные реакции на стресс, печаль и радость других людей. Используя комплексный анализ, мы обнаружили три структуры, которые показали очень разную активность в ответ на эмоции: миндалина, передняя островковая доля и височный полюс. Уровень синхронности, прослеженный в динамике от младенчества до юности, опосредовал участие в групповых взаимодействиях и гибкий эмпатический отклик социального мозга на эмоции других. Таким образом, увеличение синхронности благодаря «методу Кенгуру» позволило ранним переживаниям привязанности сформировать гибкую нервную реакцию на эмоциональные состояния других.
ВЫВОДЫ
Резильентность является основным понятием клинической науки и практики, которая до сих пор не имеет исчерпывающей биоповеденческой концепции. Два основных упущения в современных моделях резильентности связаны с ее рассмотрением исключительно в рамках нейробиологии страха и недостаточным вниманием к ее стадийному развитию. Более того, большинство моделей дают резильентности отрицательное определение (то есть отсутствие симптомов после травмы), в действительности не давая определения, что такое, собственно, резильентность.
Мы утверждаем, что для понимания резильентности необходимо принять во внимание особенности первоначального состояния млекопитающих. Детеныши млекопитающих рождаются с двумя важными ограничениями: при рождении их мозг незрел, и они поддерживают тесную близость с кормящей матерью. Таким образом, все системы, обеспечивающие резильентность, стрессоустойчивость, адаптацию и выносливость, у млекопитающих созревают под влиянием условий, которые обеспечиваются физиологией материнского тела и родительским поведением.
Мы предлагаем модель резильентности, основанную на нейробиологии аффилиации, развивающейся научной области, которая описывает нейронные, эндокринные, генетические и молекулярные процессы, лежащие в основе нашей способности формировать связи, любить, заботиться, сопереживать и принадлежать к социальным группам.
Наша модель выделяет три основных нейробиологических компонента аффилиации, которые обеспечивают резильентность. К ним относятся окситоциновая система, аффилиативный мозг и биоповеденческая синхронность.
Окситоциновая система, участвующая в процессах пластичности на клеточном, молекулярном уровнях и уровне нейрональных сетей, обеспечивает формирование привязанности, способность млекопитающих справляться с трудностями с помощью отношений и влияет на иммунную систему.
Аффилиативный мозг человека, имеющий происхождение от материнского мозга грызунов, расширился за счет включения структур более высокого порядка, которые обеспечивают эмпатию, подражание и ментализацию, и служит базой для развития других аффилиативных связей, включающих романтическую привязанность, близкую дружбу и наставничество. Он чрезвычайно пластичен, обеспечивает передачу от одного поколения к другому в ранние сензитивные периоды и формирует социально-эмоциональные компетенции.
Биоповеденческая синхронность представляет собой координацию биологических и поведенческих процессов во время социального взаимодействия, посредством этого механизма зрелый мозг матери внешне регулирует незрелый мозг ребенка и настраивает его на социальную жизнь. Биоповеденческая синхронность человека опирается на механизмы, с помощью которых у животных согласованное социальное поведение способствует разнообразию и адаптации в процессе эволюции, и развивается в рамках детско-материнской связи на основе биологических ритмов плода еще внутриутробно, на базе которых мать строит социальный невербальный «танец» в течение первых месяцев жизни. Этот синхронный обмен развивается, усложняется, превращаясь в диалог, внутри которого есть взаимность, близость и признание множества точек зрения, распространяется от взаимоотношений матери и ребенка на другие привязанности и существует на протяжении всей жизни, намечая ключевую траекторию в развитии резильентности.
Наша модель предлагает три принципа резильентности.
К ним относятся пластичность, социальность и осмысленность. В то время как первые два являются общими у человека с животными, последний специфичен для человека. Все три принципа обеспечиваются окситоциновой системой, аффилиативным мозгом и биоповеденческой синхронностью благодаря их участию в нейрональной и поведенческой пластичности, в формировании привязанности и социальности. Они также обеспечивают способность придавать смысл травме через культурные и духовные системы и аффилиативные акты, которые выходят за рамки личности.
Эта модель подтверждается лонгитудинальным исследованием трех когорт, в которых испытуемых обследовали в период от рождения/младенчества до юности. Каждая когорта соответствовала одному из типов нарушений детско-материнских отношений: связанных с матерью, ребенком или условиями среды (материнская депрессия, преждевременные роды и хроническая травма в связи с проживанием в районе военных действий), – которые оказывают долгосрочное влияние на мозг ребенка, его поведение и благополучие. В каждой когорте гипотезы были построены на конкретных исследованиях животных моделей, описывающий «недостающий компонент» (симпатия и груминг, изменчивая доступность кормовой базы и материнской близости). Мы неоднократно оценивали психическое состояние, взаимоотношения с родителями, синхронность, уровень окситоцина и гормонов стресса, когнитивные и регуляторные функции, с целью установить факторы, которые определяют резильентность у детей. В подростковом возрасте или в ранней молодости проводилось нейровизуализационное исследование социального мозга.
Нарушения в развитии возникали в разных условиях; тем не менее, они были предсказуемыми и в основном проявлялись во взаимодействиях с людьми, при этом некоторые дети демонстрировали значительную резильентность. Нейробиологические компоненты аффилиации – синхронность и система окситоцина – показали себя как факторы, способствующие резильентности в процессе развития. Эндокринная синхронность (гормональная согласованность между показателями окситоцина и гормонами стресса у матери и ребенка) проявила себя как фактор, определяющий риск дезадаптации, что свидетельствует о постоянном биологическом воздействии матери как внешнего фактора, регулирующего резильентность. В позднем детстве социальные навыки детей, обеспеченные синхронностью, выполняли роль важных показателей резильентности. Созревание регуляторных функций происходило на базе друг друга, и более высокий уровень регуляции улучшал последующее функционирование. Особое значение имели интервенции во время ранних сензитивных периодов, как, например, в результате обеспечения контакта «кожа к коже» матери с младенцем у недоношенных детей.
При нейровизуализационном исследовании социального мозга мы обнаружили изменения, определяющие резильентность. В то время как у детей, воспитанных матерями с хронической депрессией, отмечалось прерывание эмпатического ответа нервной системы, не у всех детей, растущих в травматических условиях, имелись нарушения, а только у тех, у кого синхронность с матерью была недостаточной. Нейробиологическая основа привязанности была нарушена у детей матерей, страдающих депрессией, но только у тех, у кого в последующем развилось аффективное расстройство. Аналогичным образом при нейробиологической оценке эмпатической точности недоношенные дети, имевшие достаточную синхронность с матерью, показали адекватный социальный нейронный ответ.
Таким образом, опираясь на философские и нейронаучные модели 20-го века, которые формулировали конкретный, поведенческий подход к познанию и поведению и размыли различие между мозгом и разумом, наша модель имеет целью направить внимание на системы, которые поддерживают нашу способность формировать аффилиативные связи, вступать в социальные группы и использовать отношения, чтобы справляться со стрессом, как специфические способности человека для того, чтобы противостоять, и даже процветать, перед лицом травмирующих обстоятельств.
Текущие исследования резильентности обычно фокусируются на нейробиологии стресса и страха или основаны на эпидемиологических и клинических исследованиях последствий психотравмы. В обоих случаях резильентность понимается как «отсутствие симптомов» или «сохранение психического здоровья» после перенесенных несчастий или травм4. Междисциплинарная группа5, подчеркивая необходимость смещения акцента в области психического здоровья с психопатологии на резильентность и указывая на огромное экономическое бремя и субъективные страдания, вызываемые расстройствами, связанными со стрессом, пришла к выводу, что резильентность может быть определена только постфактум травмы, когда у части людей появляются симптомы, а у части – нет.
С научной точки зрения такая позиция является проблемной. Без четкой концепции невозможно накапливать эмпирические данные и сведения о проведенном лечении.
В частности, очень важно определить, относятся ли к резильентности процессы, которые приводят к ухудшению состояния после физической или психической травмы, или те, которые однозначно способствуют увеличению силы и выносливости6,7.
Позитивная психология выделяет резильентность как ключевую составляющую благополучия8, в ее рамках запущен известный тренинг резильентности в армии США9. Также некоторые аспекты резильентности отражены в работах психоаналитиков-постфрейдистов, которые подчеркивали, что «я» является функциональным, развивающимся, существующим не изолированно, но во взаимодействии с социальной средой, в частности в трудах Sullivan10, Fromm11 и Erickson12, в работе Maslow13 о самореализации и в понятиях гуманистической психологии14. Тем не менее, эти авторы говорили не о резильентности как таковой, а о личностном росте, не интегрировали результаты систематических исследований и данные нейробиологических открытий в свои модели и концепции или даже отрицали релевантность каких-либо нейронаучных данных15,16. Специфичная для человека модель резильентности, которая, с одной стороны, учитывает его внутреннюю реальность и высшие способности, а с другой стороны, опирается на эволюционные модели и включает в свою основную концепцию результаты нейронаучных открытий, так и не была построена.
Две основные проблемы способны еще больше усложнить построение комплексной биоповеденческой модели резильентности человека. Во-первых, большинство современных усилий, направленных на понимание нейробиологических основ психических расстройств в области психиатрии, как правило, сосредоточены на особенностях, которые можно легко проверить в сравнении с другими видами. Это привело к тому, что практически все внимание сфокусировано на нейробиологии страха – нейронных, эндокринных, генетических и молекулярных процессах, которые поддерживают реакцию страха и позволяют справляться со стрессом4,17-21. Соответственно, в исследованиях часто используются межвидовые парадигмы изучения стресса, в частности механизмов, обуславливающих страх, что привело к взгляду на резильентность преимущественно с этой точки зрения22,23.
Во-вторых, не уделяется должное внимание развитию как ключевому компоненту психического здоровья, в частности резильентности, несмотря на тот факт, что все «я»-модели, по сути, являются моделями развития (то есть описывают стадийное развитие от незрелых форм к зрелым). С хорошей резильентностью не только рождаются, ее (что важно) можно воспитать. Было высказано предположение24, что для изучения резильентности мы должны обследовать детей с младенчества в течение продолжительного периода времени, чтобы выявить возрастные биологические, поведенческие и социальные маркеры, способствующие формированию у них жизнестойкости. Однако такие лонгитудинальные исследования крайне редки.
Эти две проблемы привели к довольно ограниченному, одностороннему взгляду на резильентность. Когда в дискуссии по поводу резильентности группе ведущих исследователей25 был задан вопрос «что должно быть пересмотрено», все они указали на узкое понимание резильентности, сведение ее до физиологии страха и нейробиологии стресса в теоретических и практических исследованиях как на главную проблему, заслуживающую внимания.
Новая концепция резильентности должна основываться на эволюционных механизмах, позволять проводить основательные межвидовые исследования и создавать условия для тщательного сбора данных для выявления ее специфических особенностей на разных стадиях развития, в разных условиях. С точки зрения науки чрезвычайно важно, чтобы результаты были проверяемыми (то есть открытыми для проверки уровня доказательности и фальсификации).
В дальнейшем мы предлагаем модель резильентности, основанную на нейробиологии аффилиации и предлагающую биоповеденческую эволюционную концепцию, учитывающую аспекты развития, которая не ограничена нейробиологией страха или стремлением к счастью. Наша модель учитывает фундаментальную особенность млекопитающих, чей мозг созревает как в процессе внутриутробного развития, так и в процессе воспитания, и утверждает, что созревание всех нейробиологических процессов, которые способствуют формированию резильентности, обеспечивается физиологией материнского тела и типичным для вида родительским поведением.
Кроме того, модель утверждает, что любое понимание резильентности должно учитывать первоначальную зависимость младенца от матери и огромное влияние, которое эта зависимость оказывает на структуру и функционирование мозга. Детеныши млекопитающих рождаются с незрелым мозгом, их мозг формируется под влиянием зрелой матери через физическую близость, лактацию и совокупность специфических для вида, способствующих хорошей адаптации способов ухода за детенышем, которые обеспечивают внешнюю регуляцию в системе «мать – ребенок» уникальным для системы образом26,27.
Такая внешняя регуляция незрелого мозга зрелым определяет основной механизм развития мозга у млекопитающих и обеспечивает адаптацию нейробиологической и поведенческой систем ребенка к жизни в рамках социальной экологии и ее уникальных особенностей28,29. Мы утверждаем, что процессы адаптации мозга ребенка к жизни в пределах экологической ниши и тем явным трудностям, с которыми он сталкивается, и есть сама суть резильентности, именно эти процессы определяют, что такое резильентность, и они должны стать центром внимания научных исследований.
ОСНОВНЫЕ НЕЙРОБИОЛОГИЧЕСКИЕ КОМПОНЕНТЫ АФФИЛИАЦИИ
Наша модель опирается на три основных нейробиологических компонента аффилиации: окситоциновая система, аффилиативный мозг и биоповеденческая синхронность.
Окситоциновая система
Древняя окситоциновая система, появившаяся примерно 500 миллионов лет назад, обеспечивала реакции организма на экологические проблемы, участвуя в регулировании основных жизненных функций, таких как сохранение водного и энергетического баланса, терморегуляцию в филогенетическом масштабе31,32. Таким образом, первоначально окситоцин обеспечивал выносливость, адаптацию организма к условиям внешней среды и его резильентность.
В процессе эволюции у млекопитающих окситоцин стал играть ключевую роль в родах и лактации. Тогда для млекопитающих связь между матерью и детенышем стала ключевым условием для созревания систем, которые обеспечивали совладание со стрессом32. Жизненно важные функции больше не развиваются в контексте группы, как у рыб или муравьев, а, в интимных рамках «кормящей диады», благодаря особым структурам материнского тела.
У млекопитающих окситоциновая система стала основой, обеспечивающей механизмы резильентности и аффилиации, посредством которых устойчивость, пластичность и толерантность к трудностям внешнего мира достигаются за счет социального взаимодействия, которое связывает одну особь с человеческим культурным сообществом29,33.
В целом роль окситоцина для резильентности определяется тремя компонентами, связанными с его вовлечением в нейропластичность, социальность и иммунитет.
Окситоцин задействован в процессах нейропластичности на молекулярном, клеточном уровнях и уровне нейрональной сети34-36. Нейроны могут вырабатывать окситоцин совместно с различными нейротрансмиттерами, в том числе дофамином, серотонином и опиоидами. Окситоцин вырабатывается широким спектром нейрональных клеток, таких как ГАМКергические интернейроны, глутаматергические пирамидальные клетки и другие пептидергические клетки34,37,38. Окситоцин объединяет центральные и периферические отделы нервной системы, включает массивные эпигенетические данные и, в частности, связан с переживаниями привязанности39,40. Он повышает пластичность в сети гиппокампа, повышая привлекательность объекта привязанности41, а приобретенный опыт привязанности влияет на доступность окситоциновых рецепторов42.
Пульсирующий характер высвобождения окситоцина имеет особое значение для нейропластичности, благодаря которой формируются нейробиологические системы взаимодействия с окружающей средой43. Пульсирующее высвобождение окситоцина координирует рождение при благоприятных условиях окружающей среды, знаменуя первое в жизни человека взаимодействие мозга и окружающей среды44. Всплеск окситоцина во время рождения вызывает передачу сигналов посредством гамма-аминомасляной кислоты (ГАМК), что приводит к переходу от возбуждения к торможению, тем самым координирует переход нейронов гиппокампа плода от функционирования в пренатальном к функционированию в постнатальном периоде45,46, поддерживая баланс между возбуждением и торможением на протяжении всей жизни. Оптимальный баланс возбуждения и торможения имеет решающее значение для адаптивного функционирования и обеспечивает эффект «сензитивного периода», который имеет решающее значение для устойчивости всех живых организмов47.
Окситоцин играет ключевую роль в формировании социальности. Нейронные сети, обеспечивающие формирование привязанности и связей, развивались из-за чувствительности окситоцина к повторяющимся элементам в окружающей среде, благодаря чему мать и окружение становились ценными стимулами48-50. Окситоцин в лимбических структурах побуждает младенцев предпочитать сигналы, связанные с матерью, что приводит к формированию специфической диадной привязанности51,52. В течение первых дней после рождения рецепторы окситоцина становятся ассоциированными со специфическими социальными сигналами через связи системы окситоцина с дофаминной системой вознаграждения53-55, ольфакторно-амигдалярными путями56,57, сенсорной корой54 и резким повышением отношения «сигнал/шум» пирамидальных клеток гиппокампа58. Они программируют социальное восприятие, предпочтения и память и связывают их с объектом привязанности.
Окситоцин способствует интеграции людей в социальные группы59,60. В ходе эволюции он участвовал в обеспечении социальных функций: ритмичных движениях при ухаживании у нематод61, социальных процессах у червей62, формировании пар и стай у птиц63, исключительной связи у стадных животных64 и принадлежности к группе у грызунов65, приматов66 и людей29,67. Эволюционные ограничения привели к тому, что эта гибкая, изменчивая в зависимости от внешних условий система заставляла молодых особей находиться в связи со своими родителями, функционировать в рамках их социальной экологии и включаться в социальную структуру своего вида64. Примечательно, что большая социальная поддержка и чувство принадлежности к социальной группе неоднократно ассоциировались с лучшей резильентностью68-70.
Система окситоцина у ребенка формируется в процессе ухода за ним. Исследования на животных показывают, что материнское поведение программирует доступность окситоциновых рецепторов в мозге71, а лонгитудинальные исследования на людях показывают, что окситоцин в периферической нервной системы программируется повторяющимися проявлениями заботы в детстве72-74. Окситоцин вызывает физиологическое состояние покоя, которое позволяет без страха проявлять себя в социуме и стимулирует стремление к социальному контакту через связи с дофамином в нейронах стриатума75-77. Это уникальное состояние обеспечивает основу для чувства безопасности человека, на котором может формироваться резильентность.
Наконец, окситоцин играет важную роль в функционировании иммунной системы. Исследования на людях показали связь между окситоцином и иммунными биомаркерами78,79. В клеточной культуре окситоцин снижает окислительный стресс и секрецию интерлейкина-6 (IL-6) из стимулированных макрофагов80. In vivo он снижает уровень воспалительных цитокинов, IL-6 и фактора некроза опухоли (TNF)-α81. Во время образования связей, в том числе в период становления родителем и влюбленности, и окситоцин, и IL-6, иммунный биомаркер, повышают свою активность82, а окситоцин участвует в более быстром заживлении ран83. Недавно было установлено, что кишечная бактерия, продуцирующая окситоцин (Lactobacillus reuteri), играет роль в резильентности, совладании со стрессом и более быстром заживлении ран у хозяина, что указывает не только на наличие дополнительной оси «мозг–кишечник» в выработке окситоцина, но и на значение связи «микробиом–человек» для формирования резильентности84.
Аффилиативный мозг
Термин «аффилиативный мозг» обозначает сеть взаимосвязанных структур, которые позволяют людям формировать и поддерживать близкие отношения85.
Аффилиативный мозг человека, имеющий происхождение от материнского мозга грызунов, расширился и включил в себя несколько корковых сетей высшего порядка, которые обеспечивают бессознательную мотивацию и когнитивные аспекты воспитания человека30,86. Эта глобальная нейрональная сеть, целью которой изначально являлось обеспечение ухода и связи младенца с матерью в ранние сензитивные периоды, в дальнейшем преобразовалась для поддержания социальной связи с другими людьми: любовниками, близкими друзьями и собратьями30,85.
Исследования материнского мозга на животных моделях, предпринятые в 1950-х гг., описывают основополагающую роль медиальной преоптической области гипоталамуса в подкорковой сети, которая позволяет матерям млекопитающих (и отцам у двуполовых видов) заботиться о детенышах87. Под действием окситоцина во время беременности и родов медиальная преоптическая область направляет импульсы в миндалину, что вызывает повышение осторожности у матери для обеспечения безопасности ребенка, и в вентральную область покрышки, что активирует материнскую систему вознаграждения от стимулов младенца, повышает чувствительность лимбической сети, поддерживающей материнское поведение (также включая прилежащее ядро, боковую перегородку, вентральный паллидум, ядро ложа конечной полоски и бледное ядро).
У людей эта подкорковая сеть расширилась и включает сети более высокого порядка, которые ответственны за эмпатию, подражание, ментализацию и регулирование эмоций, формируя глобальную сеть, обеспечивающую привязанность30. У 3–5% видов млекопитающих, у которых уход обеспечивают оба родителя, такая же система лежит в основе родительского поведения отца. Однако недавние исследования показали, что у матерей и отцов родительское поведение обеспечивают разные популяции нейронов88, и за отцовское, и материнское поведение отвечает одна и та же сеть, но путь к отцовству является более «когнитивным» и опирается на конкретные действия по уходу за детьми87,89.
Окситоцин играет важную роль в настройке и функционировании аффилиативного мозга. Социальное поведение людей регулируется сетью обеспечения ухода, которая содержит множество рецепторов окситоцина90. Окситоцин вызывает долгосрочную депрессию в миндалине91, чтобы ослабить ее ответ на отрицательные социальные стимулы, таким образом укрепляя сетевое соединение и обеспечивая специфичность ответа на социальные стимулы92,93.
После ослабления социального избегания окситоцин усиливает мотивацию к формированию социальных связей через перекрестные взаимодействия с дофаминовыми рецепторами в стриатуме, в частности в прилежащем ядре. Дофамин в прилежащем ядре обеспечивает целенаправленное поведение, связанное с вознаграждением, подавляет активность ГАМКергических (ингибирующих) нейронов94-97, что позволяет активировать глутаматные (возбуждающие) пути, что в свою очередь приводит к энергичным, активным, целенаправленным действиям98,99.
Оболочка прилежащего ядра содержит рецепторы окситоцина, которые образуют гетеромеры (нейроны, вырабатывающие и окситоцин, и дофамин100), и это позволяет дофаминовым нейронам, основным назначением которых является идентификация сенсорно-моторного вознаграждения, кодировать временные паттерны социального взаимодействия49,101. Это позволяет мозгу опознавать социального партнера, кодировать специфические для этой связи паттерны и получать вознаграждение от социальной синхронности96,101.
Тесные перекрестные связи «окситоцин–дофамин» позволяют гибко встраивать новую связь в личность102 и формировать сенсорно-моторные воспоминания о переживаниях привязанности103. Таким образом, в то время как дофамин обеспечивает мотивацию и активность, окситоцин обеспечивает спокойствие, необходимое для образования связи.
Эта мозговая сеть обеспечивает не только родительское поведение, но и нейронную поддержку для формирования других аффилиативных связей на протяжении всей жизни; отсюда и термин «аффилиативный мозг». В исследованиях животных104 и человека85 показано, что родительский мозг млекопитающих также участвует в поддержании парных связей у моногамных млекопитающих77, а также романтической привязанности и близких дружеских отношений у людей85. Эта аффилиативная сеть развивается в мозгу младенца в ранние сензитивные периоды благодаря целенаправленной заботе и позволяет ребенку формировать близкие отношения, влюбляться, становиться членом социальных групп от спортивной команды до наций и в конечном итоге, воспитывать своих детей.
Еще Darwin105 отмечал, что эволюционные изменения происходят во взаимодействии «родитель–детеныш», а присущая им пластичность обеспечивает появление новых форм поведения, которые со временем изменяют экспрессию генов. Соответственно, наша модель, в основе которой лежит взаимодействие «родитель–ребенок», показывает, как аффилиативный мозг использует присущую ему пластичность для формирования резильентности, выносливости и перенастройки.
Аффилиативный мозг обеспечивает резильентность несколькими способами. Оптимальная активация этой сети позволяет людям формировать и поддерживать социальные связи на протяжении всей жизни, справляться со стрессом с помощью отношений и благодаря перекрестным связям окситоцина и дофамина получать наибольшее вознаграждение от этих связей, а не из несоциальных источников (например, употребление наркотиков). Действительно, при зависимостях обнаружены нарушения взаимоотношений окситоцина и дофамина, что приводит к тому, что вознаграждение разъединяется с его социальными источниками, при этом выявлены нарушения как системы окситоцина106, так и нейропластичности107,108.
Мозг родителя формирует социальные способности ребенка. Мы обнаружили, что активность мозга родителя в младенчестве позволяла прогнозировать способность к регуляции эмоций, совладанию со стрессом и формирование симптомов у ребенка в течение первых семи лет жизни109-111. Вместе с тем заботливое и отзывчивое воспитание в перспективе обеспечивало формирование аффилиативного мозга в подростковом возрасте112,113. Наконец, большая ассоциативная кора человеческого мозга позволяет обрести смысл через любовь к абстрактным идеям, таким как родина или Бог, и распространять привязанность на других людей, домашних животных, флору и фауну Земли, и все это обеспечивает одна и та же сеть85.
Биоповеденческая синхронность
Биоповеденческая синхронность является основным механизмом, обеспечивающим социальность и аффилиацию у человека. Она определяется как «координация биологических и поведенческих сигналов между социальными партнерами в момент социального контакта» и описывает механизм, с помощью которого зрелый мозг родителя внешне регулирует незрелый мозг младенца и настраивает его на социальную жизнь29,114,115.
Биоповеденческая синхронность создает шаблон для координации биологического с социальным и психическим, делает возможным слияние автономного «я» с автономным «другим» и объединение моментов межличностного соответствия с моментами несоответствия, моментов разлуки и встречи – все в рамках безопасного диалога.
В многочисленных исследованиях, охватывающих период от младенчества до взрослой жизни среди широкого круга здоровых и групп населения высокого риска в различных культурах, мы показали, что эти «драгоценные социальные моменты», когда родитель и ребенок координируют свое невербальное поведение, представляют собой моменты биологической координации. Например, только во время этих эпизодов наблюдалась синхронность между сердечными ритмами матери и младенца116, координированный выброс окситоцина117 и синхронизация социального мозга обоих118.
Синхронность сочетается с лучшим совладанием со стрессом73, более высокой дыхательной синусовой аритмией119 и лучшим функционированием иммуннитета120, открывая механизм, посредством которого скоординированное социальное поведение снижает стресс и улучшает резильентность.
Связь поведенческой и биологической синхронности берет свое начало в утробе матери121, когда биологические ритмы младенца включаются в социальный диалог, который превращает биологическое в отношенческое, а индивидуальное – в межличностное. Паттерны невербальной синхронности проявляются в диадических отношениях постоянно, расширяясь в символической и межличностной сложности122, и, как предполагает теория динамических систем123, такое расширение репертуара с опорой на первичный опыт отражает механизм резильентности. Примечательно, что все формы физиологической синхронности (нейронная, эндокринная и автономная) встроены в поведенческую координацию, что подтверждает нашу основную гипотезу о том, что поведенческая синхронность формирует физиологическую связь и что резильентность базируется на поведении29,124,125.
Биоповеденческая синхронность, наблюдаемая в первые месяцы жизни, представляет собой крайне важный опыт, полученный в сензитивный период, который позволяет прогнозировать множество компонентов, связанных с резильентностью от рождения до юности, в том числе регуляцию эмоций, символическую компетентность, способность справляться со стрессом, выраженность экстернализированных и интернализированных симптомов, а также социальные развитие мозга30,114,126,127.
В процессе развития сопоставление ребенком невербальных аффективных реакций превращается в обоюдный обмен, который порождает, как брошенный камень порождает круги на воде, растущие символические, лингвистические и социальные компетенции ребенка, развивается эмпатия, мировоззрение и близость, все они построены на ритмичном невербальном ядре и служат резильентности (см. ниже). Это перекликается с представлениями Maslow13 о том, что «я» включает в себя как то, чем является человек, так и то, кем он может стать. В то же время, будучи специфическим для человека механизмом, который развивается в течение всей человеческой жизни, биоповеденческая синхронность опирается на длинную эволюционную линию необходимых для выживания социальных механизмов млекопитающих и других эусоциальных (гиперсоциальных) видов, обеспечивающих выносливость и резильентность.
В ходе эволюции, от бактерий до человека, синхронность основывается на процессах, которые связывают два организма (или две сущности) в единую биологию. Последние достижения в области квантовой физики позволяют предположить, что такое взаимодействие началось еще до появления жизни, что можно увидеть в феномене «квантовой запутанности», при котором связь частиц во времени и пространстве, объединяющая две единицы вместе, придает их объединению неизмеримую силу и стойкость.
ТРИ ПРИНЦИПА РЕЗИЛЬЕНТНОСТИ
Учитывая вышеупомянутые основы аффилиативной нейробиологии (окситоциновая система, аффилиативный мозг и биоповеденческая синхронность), наша модель выделяет три принципа, определяющих, что такое резильентность. Несмотря на то, что все три элемента необходимы для формирования жизнестойкости человека, они существуют в различных комбинациях у разных людей в разных культурах и выражаются по-разному в зависимости от возраста и стадии (Рисунок 1).
Резильентность включает в себя пластичность
Прежде всего, резильентность включает в себя механизмы, которые способствуют гибкой адаптации к изменяющимся условиям, изобретательному использованию актуальных условий для личностного роста и способности упорно достигать долгосрочных целей в сочетании со способностями к изменению и поднастройке. То есть резильентность подразумевает пластичность.
Пластичность основывается на нейробиологических сетях, которые обеспечивают социальную адаптацию, физическую выносливость и устойчивость за счет гибкого приспосабливания к различным условиям128,129. По-видимому, наиболее пластичным формирование связей является у млекопитающих. Большая нейропластичность привела к развитию живорождения (внутренней беременности) и физиологической реорганизации системы «мать–ребенок», что сделало возможным созревание плода в материнском теле130. Также огромная нейропластичность необходима для того, чтобы сделать новорожденного наиболее значимым объектом для матери по сравнению со всем остальным131.
Как уже было отмечено, окситоциновая система играет ключевую роль в нейропластичности, имеющей решающее значение для формирования привязанности, а послеродовой период соответствует наибольшей пластичности мозга взрослого человека132.
Компонент резильентности «пластичность» включает в себя две особенности: а) резильентность является интегративной и регулирующей; б) резильентность связана со временем.
Резильентность является интегративной и регулирующей
Регуляция способствует гибкой интеграции компонентов системы в функциональное целое, формируя «я», индивидуальность, свободу действий и благополучие посредством формирования новых, специфичных для человека, специфичных для диады и специфичных для культуры конфигураций. Многие исследования проблем развития были направлены на понятие «регуляции», причем некоторые полагают, что это наиболее важная концепция в понимании нарушений развития133,134.
Во многих областях «регуляция» имеет системный уровень. Он определяет, как различные компоненты системы динамически объединяются в функциональное целое, как высшие и низшие элементы иерархически организуются во времени и как компоненты внутри системы интегрируются в режиме реального времени с компонентами ближайшего окружения123,135-137. Концептуальные модели предполагают, что созревание регуляторных процессов происходит от биологических к эмоциональным, затем к управлению вниманием и, в завершение, к саморегуляции138, а совместная регуляция «родитель–ребенок» (синхронность) обеспечивает созревание регуляторных навыков более высокого порядка, таких как управление вниманием и самоконтроль139-141.
Резильентность связана со временем
Резильентность ограничена во времени, имеет процессуальный характер и развивается от простого к сложному, от биологического к психическому. «Временной» компонент резильентности имеет решающее значение не только в процессе эволюции (в филогенезе) и от младенчества до взрослой жизни (в онтогенезе), но и на уровне конкретного социального опыта.
Социальные контакты всегда разворачиваются в рамках временного промежутка, когда два или более участников создают новый «танец» из совпадений и несовпадений, которые определяют поведение, физиологические реакции и психическое состояние участников. Временный характер этих встреч позволяет формировать новые формы из существующих единиц. Следовательно, время является неотъемлемым компонентом резильентности (способности к перенастройке), и это отражается в «синхронности», конструкте, связанной со временем.
Резильентность является социальной
Социальность лежит в основе выживания и адаптации, а виды, которые лучше используют социальные механизмы для координированных действий, имеют значительное преимущество в выживании. Это элегантно описано энтомологом E. Wilson142 в работе «Социальное завоевание земли» («The social conquest of earth»), где он утверждает, что люди достигли превосходства среди позвоночных, а муравьи – среди беспозвоночных, с точки зрения численности населения, распространения по всей земле и выносливости, благодаря их эусоциальности (гиперсоциальности), которая включает в себя способность к совместным действиям членов группы и передачу социальной организации от поколения к поколению.
Примитивные механизмы синхронности обнаруживаются у муравьев, рыб и птиц, при этом координация биологических и поведенческих процессов обеспечивается вазотоцином, родительской молекулой окситоцина и вазопрессина млекопитающих32,62,143. Таким образом, биоповеденческая синхронность у человека опирается на долгую историю социальных механизмов, которые способствовали резильентности путем координации действий. В соответствии с поведенческим принципом аффилиативной нейробиологии эти механизмы прошли отбор с акцентом на поведение: социальное поведение в группе у видов, не относящих к млекопитающим, и аффилиативные связи у млекопитающих. Однако примечательно, что, хотя одиночество опасно для благополучия любого живого организма144, «социальный» компонент резильентности сильно варьируется, и наблюдается широкая изменчивость по всему животному царству, что сочетается с большой изменчивостью плотности и локализации рецепторов окситоцина145,146.
Социальная моногамия
Социальная моногамия знаменует собой первое расширение связи «мать–ребенок» до других привязанностей в семье, в частности, формирования пары и отцовства. Исследования социальной моногамии проводились у нескольких видов приматов (тамарины, мартышки и лемуры) и пяти видов грызунов, происходящих из одной линии (степные полевки, китайские полевки, калифорнийские мыши, карликовые хомяки Кэмпбелла и монгольские песчанки)148.
Моногамия обеспечивает основу для отцовства. Целенаправленное родительское поведение встречается в основном у социально-моногамных видов149. У них отцовство развивается параллельно с материнским уходом, а родители координируют свою заботу по отношению друг к другу150. Участие отца улучшает резильентность молодых млекопитающих, повышает выживаемость потомства, увеличивает размер помета и темпы роста151-156.
Хотя конкретные экологические причины, которые привели к формированию модели совместного воспитания с участием двух родителей у людей, неизвестны, участие отца в воспитании стабилизировало моногамные партнерские системы. После того, как социальная моногамия устанавливается у вида, она способствует появлению сложных социальных форм поведения, которые улучшают резильентность154,157.
Как отцовство, так и парные связи представляют собой расширение связи «мать–младенец», перепрофилирование одних и тех же нейронных сетей и молекулярных процессов и первое проявление последовательной и многообразной нейробиологии аффилиации. Моногамия также требует координации трех внутрисемейных привязанностей (материнство, отцовство и парная связь) в семейную единицу, и такая координация проложила эволюционный путь к человеческой семье и в конечном итоге сложной социально-культурной организации, что обеспечило наилучшую резильентность людей в животном мире.
У людей участие отца в воспитании обеспечивает значительную резильентность. На протяжении всей истории человечества отцы были основным источником косвенной помощи, контролировали материальные ресурсы, физические условия и социальный статус, в которых развивались дети158,159. Исторические отчеты указывают на тесную связь между наличием родителей и детской смертностью в доиндустриальных США и Европе160, а антропологические исследования показывают, что мужчины с большим количеством земли или более высоким социальным статусом демонстрируют больший репродуктивный успех161,162.
В современных обществах более активное участие отца в воспитании повышает резильентность детей, улучшает их психическое здоровье, способствует более высоким академическим и профессиональным достижениям, а также улучшает способность к саморегуляции163,164. Дети, чьи отцы участвовали в воспитании, менее агрессивны и склонны разрешать конфликты, сохраняя уважение, и вести диалог165, а эпидемиологические исследования показывают, что дети, росшие без отца, более склонны к агрессии, нарушению закона и проблемному поведению166,167.
Сложные социальные организации
Хотя социальная моногамия представляет собой первое расширение связи между матерью и ребенком до семейной единицы, считается, что сложная иерархическая социальная организация развивается только у гомининов и растет параллельно с увеличением размера мозга168. Недавние исследования западных горилл обнаружили иерархическую социальную модульность, определяющую не только сложное аффилиативное поведение в рамках обширных родственных групп, но также взаимное сотрудничество между неродственными группами в рамках сезонных коалиций, имеющих общую цель, аналогичных структуре маленькой деревни у людей169.
Такие организации обеспечивают совместный сбор широко рассредоточенных продуктов питания и защиту от хищников, повышая резильентность благодаря совместным действиям вне семьи. Среди приматов, живущих в группах, таких как шимпанзе, наблюдается постконфликтное примирение, которое позволяет членам группы полюбовно разрешать конфликты и поддерживать социальные связи, и эти действия, следующие после акта агрессии, приводят к увеличению окситоцина в моче170.
Исследование на мартышках показало, что чем крепче связь между аффилиативной парой (того же или противоположного пола), измеряемой с точки зрения длительности отношений, времени, проведенного вместе, и проявлений аффилиативного поведения, тем выше эндокринная синхронность колебаний окситоцина в моче171, что указывает на наличие у приматов биоповеденческой синхронности, предшествующей таковой у людей.
Биоповеденческая синхронность – механизм, специфичный для человека
Основываясь на этих социальных механизмах, которые обеспечивают совладание со стрессом, групповую сплоченность и сенсорно-моторную координацию, биоповеденческая синхронность представляет собой специфичный для человека механизм, посредством которого два человека могут взаимно влиять на физиологию друг друга без физического контакта, посредством координации лицевых аффективных сигналов, которой нет у других приматов и грызунов29.
У человека синхронность развивается в течение всей жизни и постепенно усложняется, включает совместное построение общего нарратива, способность принимать различные точки зрения и способность сопереживать боли, действиям, эмоциям и психическому состоянию другого человека. Развитие синхронности начинается с распознавания матерью биологических ритмов младенца в своей утробе и заканчивается построением близких отношений между взрослыми, наполненных взаимной заботой.
Резильентность включает в себя осмысленность
В то время как первые два принципа резильентности основаны на механизмах, общих для всех видов, смыслообразущий компонент является специфичным для человека. Для формирования концепции резильентности у человека мы должны объединить общие для всех видов механизмы выносливости, многообразия, адаптации и совладания со стрессом со способностью человека придавать значение трудностям, невзгодам и травмам.
Люди способны придавать смысл травме, используя коллективные культурные или религиозные мифы, или в других случаях, наделяя персональными смыслами особые действия, обычно такие, которые ведут к укреплению аффилиативных связей, или акты альтруизма, выходящие за пределы личности.
Во многих исследованиях подчеркивается роль духовности в способности восстанавливаться после перенесенных невзгод или использовать травму для роста172-174. Исследования также указывают на важность щедрости для резильентности175 и на утешительную функцию религиозных сообществ, которые придают коллективный смысл, а также оказывают поддержку176,177. W. James в работе «Воля к вере»178 («The will to believe») рассматривал веру как преднамеренный выбор, который дает опору и позволяет человеку сформировать личностно-смысловой взгляд на реальность, придавать значение травме и трудностям. Его знаменитая метафора превращения дискретного опыта в осмысленное целое – «живые электрические провода», которые светятся и сияют, в отличие от «мертвых проводов», которые остаются расплывчатыми и тусклыми, – элегантно описывает эту функцию веры, способствующую резильентности.
Создание смысла вводит дименсию «будущее» в концепцию резильентности, расширяя временной горизонт за пределы «помнящего присутствия»179 других приматов. Это подчеркивает целенаправленную функцию культурных мифов, которые создают люди и которые выходят за рамки жизни человека и подпитывают внутренние резервы резильентности перед лицом трудностей.
Обретение смыслов, выходящих за рамки индивидуальной жизни, не только является ключевой особенностью резильентности, но и находит объяснение в двух нейробиологический системах аффилиации. Carter76 предположил, что окситоциновая система обеспечивает нейробиологический субстрат для духовности благодаря своей роли в поддержании любви, заботы, эмпатии и морального подъема, в частности потому что позволяет млекопитающим испытывать «состояние бдительности без страха», что значит «быть полностью в настоящем моменте без ощущения потенциальной опасности». Точно так же нейронные структуры, связанные с «аффилиативным мозгом» и формирующиеся в ранние сензитивные периоды, позволяют людям распространять любовь на незнакомцев, социальные группы и абстрактные идеи, проявлять великодушие за пределами непосредственных связей индивида.
Тем не менее сильные культурные мифы разных поколений, системы смыслов и религиозные верования способны нивелировать первый принцип резильентности – гибкость – путем ужесточения обычаев, обязательств, требуя подчинения, ужесточая контроль. Такие группы часто функционируют благодаря тесной внутренней сплоченности, достигаемой за счет усиления нейронной и поведенческой синхронности между ее членами до гиперсоциального уровня перед лицом реальной или мнимой опасности. Например, на протяжении всей истории человечества солдаты проходят интенсивную подготовку к скоординированным действиям, и эта моторная синхронность позволяет устранить когнитивную эмпатию во время битвы, чтобы сражаться и уничтожать тех, кто к группе не относится. Социальный принцип резильентности становится более значимым для группы и неприменим к тем, кто вне ее.
Примечательно, что и окситоцин, и нейрональная синхронность задействованы в таком разделении «относящийся к группе / не относящийся к группе», построенном на древних механизмах, позволяющих немедленно отличить друга от врага, чтобы защитить близких. К примеру, мы изучали нейронную реакцию израильской и палестинской молодежи с использованием магнитоэнцефалографии (МЭГ), когда они наблюдали за тем, как переносили боль представители их группы и те, кто к ней не относился. В течение первых 500 мс, представляющих автоматический ответ мозга на чужую боль, молодые люди реагировали на боль и «своих», и «чужих». Однако спустя эти полсекунды сопереживания высшие процессы блокировали естественную эмпатическую реакцию мозга на боль представителя другой группы, давая проявиться только реакции на боль представителя своей группы180.
Два процесса помогали отключить эволюционно-древнюю эмпатическую реакцию на конспецифическое состояние при дистрессе: повышение уровня окситоцина и усиление нейронной синхронизации между членами группы. Таким образом, окситоцин и нейронная синхронизация определялись системой смыслов, которая подавляла эмпатию к тому, кто не был членом группы.
Исследование роли окситоцина в отвержении «чужих»181, ставит вопрос, как интегрировать роль систем смыслов, которые, способны, с одной стороны, улучшать резильентность путем создания сообществ и придания травме межпоколенческого значения, а с другой – вызывать агрессию и предубеждение к тем, кто не относится к группе. Возможно, одно из решений заключается в том, чтобы направлять постоянные усилия на то, чтобы придать древним системам смыслов гибкость и человечность, чтобы старые ритуалы не становились ригидными и распространялись на всех людей.
СИНХРОННОСТЬ ОТ МЛАДЕНЧЕСТВА
К ЗРЕЛОСТИ: РАЗВИТИЕ РЕЗИЛЬЕНТНОСТИ
Созревание синхронности происходит не только в ходе эволюции животных, но и на протяжении всей человеческой жизни. Первоначально синхронность проявляется главным образом в отношениях матери и ребенка, первичной связи у млекопитающих, а затем распространяется на другие социальные связи, включая отца, наставников, близких друзей и романтических партнеров, все человечество, проявляется в чувстве синхронности с природой, искусством и священным опытом.
Эти представления предоставляют биологические и научные доказательства концепции Winnicott, изложенной в «Игре и реальности»182 («Playing and Reality»), о непрестанном присутствии матери в качестве основы для формирования символов, игры, творчества и духовного опыта. Синхронность усложняется по мере разнообразия репертуара, перехода к символическому уровню и зависит от степени взаимности в детстве и подростковом возрасте, настраивает социальный мозг по мере получения нового опыта на понимание других, проявление сочувствия к чужим страданиям и участие в отношениях183. Корни синхронности происходят из эволюционно древних паттернов, в биологических ритмах плода эти переживания проявляются на физическом плане, что позволяет резонировать всей истории отношений в моменте встречи человека с человеком.
В то время как философские взгляды на «объединение» предполагают, что «я» строится из микроидентификаций, которые раскрываются во время конкретного повседневного опыта, синхронность добавляет элемент, который «я» берет из конкретных паттернов с участием значимого человека. Наша модель рассматривает подробнее развитие этого феномена как в ходе эволюции, так и на протяжении жизни человека, и показывает его вклад в формирование устойчивости к неблагоприятным условиям.
Синхронность между матерью и младенцем начинается, когда мать распознает первые биологические ритмы младенца в утробе, такие как сердечные ритмы и циклы сна и бодрствования, которые посылают сигналы к плаценте и материнскому мозгу184-186. После рождения матери вовлекают эти знакомые ритмы в диадический обмен.
Исследования 1970-х годов показали, как взаимодействие между матерью и младенцем «лицом к лицу» строится на модели «действие–пауза» повторяющихся биологических актов, таких как сосание или плач187,188. От простого участия в этих биологических ритмах синхронность постепенно развивается в эмпатические отношения между взрослыми, в рамках которых возможен диалог.
Мы изучали проявления синхронности между матерью и ребенком от рождения до 25 лет и наблюдали, как при взаимодействии в течение четверти века сохранялись одни и те же невербальные ритмические паттерны, колебания возбуждения и положительные пики. Например, в некоторых диадах уровень возбуждения постоянно колебался между низким и средним, в то время как в других взаимодействие происходило в виде быстрых пиков положительного возбуждения. Такая стабильность создает базовый порядок, пока более сложные и творческие паттерны встраиваются в диалог и образуют знакомое и единое событие.
Помимо обеспечения «ритма безопасности» для улучшения резильентности особенно важны две дополнительные функции синхронности. Во-первых, микроструктура синхронного опыта – это постоянное чередование разрыва контакта и его восстановления. Согласно Tronick189 матери синхронизируются с младенцем только около 30% времени; таким образом, диады проводят больше времени в неверно скоординированных состояниях, чередующихся с драгоценными моментами синхронности. Сторонники психоанализа и теории развития189,190 подчеркивают важность таких циклов соответствия и несоответствия для обучения младенцев тому, как переживать моменты отсутствия сонастройки и как устранять недопонимание, присущее человеческому диалогу.
Описаны два типа отклонений от паттерна «длительное несоответствие–короткое соответствие». Первое – гиперсинхрония, обнаруживается у тревожных матерей и выражается в увеличении числа эпизодов соответствия (более 45% времени); второе – абстиненция, наблюдаемая у депрессивных матерей, связана с почти полным отсутствием сонастройки. Оба приводят к трудностям саморегуляции у младенцев191,192. Таким образом, синхронность, создает серию микрособытий, состоящих из постоянного чередования разрыва контакта и его восстановления, и в безопасных условиях учит детей переносить фрустрацию. В возрасте около 9 месяцев у младенцев появляется способность самостоятельно восстанавливать контакт193, что готовит их к построению равных отношений с друзьями и партнерами.
Второй функцией синхронности, способствующей резильентности, является ее роль в качестве первого контекста для развития прогностических способностей мозга. Недавние модели «прогнозирующего кодирования» рассматривают мозг как вычислительное устройство, главная роль которого заключается в повышении адаптации путем минимизации энтропии и повышения уверенности. Нейронные волны играют важную роль в кодировании с построением прогноза: альфа-волны участвуют в построении прогноза, бета-волны – в оценке его точности, а гамма-волны – в оценке ошибки прогноза, постоянном точечном соотнесении прогноза с поступающей информацией196,197. Опыт синхронности обеспечивает шаблон для полиритмической когерентности, который создает мультисенсорный образ тела в мире197 и включает в себя интеграцию альфа-, бета- и гамма-ритмов при формировании социальных прогнозов в реальном времени112.
Используя экологическую парадигму и методы гиперсканирования, мы обнаружили связь гамма-ритмов между мозгом матери и ребенка118, романтических партнеров198 в моменты поведенческой синхронности. В исследованиях как у животных199,200, так и у людей201,202 гамма-ритмы показывают уровень зрелости мозга, что указывает на роль синхронности в тонкой настройке этой зрелости. Гамма-ритмы и ошибки прогноза в висцеромоторной коре и областях, ответственных за мотивацию, усиливают чувства, но из-за неоднородности этих областей различия между «я» и другими стирается203. Таким образом, изучение синхронности может помочь по-новому взглянуть на социальное созревание мозга в реальных условиях.
Стадии развития синхронности
По результатам многочисленных продольных и поперечных исследований мы выявили пять различных этапов развития синхронности от периода беременности до юности и показали наличие индивидуальной стабильности между этими стадиями и ее чувствительность к определенным неблагоприятным условиям.
Подготовка к синхронности связана с постепенным знакомством матери с биологическими ритмами плода: циклами сна–бодрствования, консолидирующимися примерно на 31–32-й неделе беременности204,205, за которым следует организация сердечных ритмов примерно на 33-й неделе беременности206,207. Эти циклы согласуются с реакцией плаценты208, и хорошая организация этих биологических ритмов прогнозирует лучшую синхронность между матерью и младенцем в 3 месяца121.
Период новорожденности: поведение матери в послеродовом периоде
Сразу после рождения и в течение первых 6 недель жизни человеческие матери, как матери всех млекопитающих, проявляют видоспецифичный репертуар материнского поведения, который у людей включает в себя рассматривание лица и тела младенца, выражение положительных эмоций, высокие вокализации «материнского языка» и нежные прикосновения. Однако в отличие от других млекопитающих у людей матери координируют свое поведение с краткими моментами настороженности новорожденного. Таким образом, в норме при рождении у младенца происходит координация между его внутренним состоянием и реакцией социального мира.
Материнское послеродовое поведение в неонатальном периоде обеспечивает основу для развития символической компетентности209, лучшего когнитивного развития и меньшего количества экстернализированных и интернализированных симптомов в раннем детстве210 и коррелирует с уровнем окситоцина у родителей211.
Младенчество: аффективная синхронность
С третьего месяца жизни мать и младенец начинают участвовать в интерактивном «танце», создают специфичный для диады ритмичный диалог, используя взгляды, выражение эмоций, ко-вокализации и прикосновения. Этот невербальный опыт играет ключевую роль в социальном, эмоциональном, когнитивном развитии ребенка114. Матери и отцы участвуют в специфичных для родителей формах синхронности, более ритмичных у матерей и объект-ориентированных у отцов212.
Аффективная синхронность между родителями и детьми связана со множеством гормонов, питающих привязанность, таких как окситоцин, вазопрессин, бета-эндорфин, пролактин, кортизол и слюнная альфа-амилаза, а также с иммунными биомаркерами, включая IgA и IL-6 в слюне213. Помимо этого, она связана с активацией аффилиативного мозга у матерей214 и отцов89. Невербальная синхронность также обнаруживается при взаимодействии триады «мать–отец–ребенок»215, что создает основу для социального участия детей в отношениях внутри группы и культуры.
Преддошкольники/дошкольники: символические игры и совместное конструирование воображаемых нарративов
На втором и третьем годах жизни дети начинают участвовать в символической игре и наполнять объекты символическим значением и «похожими на рассказ» символическими последовательностями. Синхронность с матерью и отцом в младенчестве216 не только определяет уровень сложности символов, также организацию символических игр переопределяет временной контур ритмического обмена младенца с матерью и отцом – коротких, случайных и многочисленных интеракций с отцом и более редких и длительно выстраиваемых – с матерью126.
В дошкольные годы дети начинают совместно строить диалог, используя будущее и прошлое время, воображаемые сценарии и альтернативную реальность, в которую они могут погрузиться. Эти игровые творческие способности опираются на невербальную синхронность первых месяцев жизни210 и превращают синхронизированный диалог в социальное событие, включающее в себя творчество, использование языка и деятельность формирующегося сознания, которые выражают внутреннюю реальность. Взаимодействие дошкольников с матерью и отцом предопределяет умственные способности детей и развитие нравственной позиции в детстве и подростковом возрасте114.
На этом этапе у детей появляются «лучшие друзья», и они вступают в социальные институты. Опыт аффективной синхронности формирует социальные компетенции ребенка со сверстниками в зависимости от культурных традиций217. Уровень родительского окситоцина, гены OXTR и ранняя синхронность позволяют прогнозировать синхронность детей с их первым лучшим другом72.
Позднее детство / подростковый возраст: эмпатический диалог
Начиная примерно с 9–10 лет и вплоть до подросткового возраста, дети заметно сокращают количество «игровых» взаимодействий со своими родителями, и диалог приобретает словесную форму – вид взаимодействия, который предполагает разрешение конфликтов, обмен информацией и, в норме, обсуждение ребенка с родителями пережитого опыта, идей, чувств, обмен мнениями и планами на будущее.
Синхронизированный диалог на этом этапе запускает у ребенка способность к поведенческой, эмоциональной и когнитивной эмпатии, умение планировать заранее, разрабатывать, сотрудничать, проявлять мотивацию и понимать точку зрения другого человека. Такие социальные способности, особенно на этой стадии, когда фокус привязанности переносится с родителей на друзей, имеют решающее значение для благополучия детей и связаны с устойчивостью перед лицом трудностей и созреванием социального мозга113,218-220.
Взрослая жизнь: взаимность, близость и взаимопонимание
Если связь между матерью и ребенком была «достаточно хорошей» и синхронность развивалась в соответствии с возрастными нормами, создавая пространство как для согласованного контакта, так и для фрустрации, мать и выросший ребенок могут встретиться друг с другом как двое взрослых, которые все еще сохраняют свои роли, но могут вести диалог с уважением к зрелости друг друга, который будет отражать их отношения в целом. Он опирается на ранние привычные ритмы и перекликается со всеми этапами развития, но этот диалог, который является взаимным и уважительным, интимным и автономным, знакомым и безопасным, все же отличается от диалога с партнером.
Такая надежная синхронность позволяет людям вступать в другие взаимные и доверительные отношения и перекидывает мост к следующему поколению, которое может превзойти родителя благодаря способности повзрослевшего ребенка пробуждать в своем собственном мозгу диадические переживания от взаимодействия с родителем в его отсутствие.
В целом синхронность, которая постепенно обогащает социальный репертуар младенца, обеспечивает созревание более сложных умственных способностей, открывает путь изменчивости при сохранении порядка, разнообразию при сохранении стабильности. Синхронность касается той области жизни, где биологическое объединяется с социальным, чтобы придать смысл, сформировать узы и противостоять трудностям.
ВОСПИТАНИЕ РЕБЕНКА С ХОРОШЕЙ РЕЗИЛЬЕНТНОСТЬЮ: ЛОНГИТУДИНАЛЬНОЕ ИССЛЕДОВАНИЕ ТРЕХ КОГОРТ ВЫСОКОГО РИСКА
Наша модель предполагает, что биологические и социальные положения, заложенные в связи между матерью и ребенком, обеспечивают основу для резильентности на протяжении всей жизни. Однако у многих детей по всему миру эти положения оказываются нарушены. Чтобы улучшить понимание резильентности, мы должны рассмотреть неблагоприятные факторы в отдельности, изучить, что именно нарушается в каждом отдельном случае, и проверить, как эти упущения влияют на результат.
Мы предположили следующее: исследования на людях должны начинаться с рождения или сразу после него; предпочтителен лонгитудинальный дизайн; изучение «недостающего компонента» должно строиться с учетом данных конкретных исследований на животных моделях, выявивших эти положения, путем оценки их последствий для детского мозга и поведения30.
Существуют три основных источника нарушений детско-материнских отношений: связанные с матерью, ребенком и условиями среды. Каждый из них затрагивает миллионы детей во всем мире.
Послеродовая депрессия отмечается у 15–18% рожениц в индустриальных обществах и достигает 30% в развивающихся странах221. Мы предположили, что работа Meany222 о долгосрочном влиянии недостаточного ухода (в виде вылизывания и груминга) у крыс на выработку окситоцина и реакцию на стресс у детенышей может дать представление о долгосрочных последствиях материнской депрессии.
Преждевременные роды происходят в 10,5% живорождений в индустриальных обществах223, и хорошо известен их негативный эффект, в частности связанный с разлукой с матерью в период выхаживания и влиянием на системы жизнеобеспечения, что согласуется с моделью «материнской близости» Hofer27.
Стресс на ранних этапах жизни имеет долгосрочные негативные последствия для развития. Каждый пятый ребенок в мире растет в условиях хаоса, иммиграции, отсутствия продовольственной безопасности или жилья, племенной или этнической войны, бедности и насилия. Животная модель, которую можно считать аналогичной, предполагает «варьирование доступности кормовой базы»224,225 у индийских макак, в которой матерям пища то доступна, то недоступна или труднодоступна, и таким образом для детеныша непредсказуемым образом меняются периоды, когда мать заботиться о нем, и периоды, когда забота сводится к минимуму. Установлено, что такие условия оказывают худшее влияние на потомство – с точки зрения развития мозга, реакции на стресс и поведения – по сравнению с постоянными хорошими или плохими условиями жизни, что позволяет предположить, что непостоянство условий в раннем периоде жизни является наиболее вредным для резильентности детей.
Чтобы понять резильентность с точки зрения нейробиологии развития, мы исследовали три когорты матерей и младенцев в период от рождения (или младенчества) до подросткового возраста / ранней молодости, уделяя особое внимание тому, как нейробиологические компоненты аффилиации отличались у детей в зависимости от жизненного пути. В каждой группе имелось одно из вышеупомянутых нарушений детско-материнской связи, и гипотезы основывались на аналогичных животных моделях.
В когорту послеродовой депрессии были включены дети, матери которых страдали хронической депрессией в течение первых лет жизни ребенка. Когорта, живущих в условиях войны, включала матерей и детей, живущих в зоне непрерывных боевых действий, а группа недоношенных включала неврологически благополучных недоношенных детей, у половины из которых в неонатальном периоде был обеспечен контакт «кожа к коже» («метод Кенгуру»).
В течение всего детства проводилась повторная оценка показателей синхронности, навыков саморегуляции, уровня окситоцина, гормонов стресса, уровня психического развития и в конечной точке – нейровизуализационное исследование социального мозга.
Послеродовая депрессия
В нашу когорту были включены только матери, физически здоровые, имеющие партнера, старше 21 года, чей доход был выше черты бедности, чтобы отделить проявления собственно депрессии от часто сопутствующих негативных факторов (матери-одиночки, матери-подростки, бедность). Оценка депрессии проводилась неоднократно в течение первого года и снова через 6 и 10 лет. Мы сформировали две когорты: дети матерей, которые страдали хронической депрессией от рождения до достижения ребенком 6 лет, и здоровый контроль.
Послеродовая депрессия у матери увеличивает риск психических заболеваний
Наличие депрессии у матери заметно повышает вероятность психических расстройств у детей, даже в семьях с низким риском. В возрасте шести лет у 60% детей матерей, у которых было диагностировано большое депрессивное расстройство как в 9 месяцев, так и в 6 лет, и которые сообщали о том, что в целом депрессия наблюдалась в первые годы жизни ребенка, был установлен психиатрический диагноз Оси I (по сравнению с 15% в контрольной группе), причем наиболее распространенными расстройствами были тревожное расстройство и расстройства поведения226. В 10 лет и в подростковом возрасте более, чем у 50% этих детей все еще сохранялся психиатрический диагноз, даже когда у матери отмечалась ремиссия, что указывает на долгосрочный эффект такого раннего воздействия. Выраженность экстернализированых и интернализированных симптомов была больше у детей матерей, страдающих депрессией227.
Синхронность способствует резильентости
Матери, страдающие депрессией, не могли обеспечить ребенку соответствующий уход, необходимый для развития. В 9 месяцев микроанализ невербального поведения показал у матерей с депрессией минимальные показатели зрительного контакта, положительного аффекта и нежных прикосновений и минимальную синхронизацию со своим ребенком191. Поскольку синхронность развивается с течением времени, матери, страдающие депрессией, не могли развить более зрелые формы взаимного диалога.
Синхронность была индивидуально стабильной от рождения до подросткового возраста, и более низкая синхронность у детей матерей с депрессией сочеталась с развитием психических заболеваний и социальной отгороженностью у детей. В 6 лет дети матерей, страдающих депрессией, демонстрировали недостаточную поведенческую эмпатию228. В 10 лет они демонстрировали более низкие исполнительские функции и недостаточное распознавание эмоций. Более низкая синхронность предопределяла снижение показателей социально-эмоционального интеллекта.
В то же время синхронность функционировала как компонент резильентности. Если дети матерей с депрессией все же получали больше синхронности (от своих отцов, из-за большей активности системы окситоцина или из-за врожденной общительности ребенка), это служило защитным фактором. Способность детей функционировать более адекватно в социальном мире, создавать дружеские отношения и участвовать во взаимодействиях со сверстниками, опосредованных синхронностью, заметно снижает риск психических расстройств, уменьшает влияние материнской депрессии в ранние годы ребенка на его исполнительские способности и распознавание эмоций. Этот эффект был особенно заметен в позднем детстве, в период, когда отношения между сверстниками начинают оказывать большее влияние на жизнь детей. Это подтверждает наше утверждение, что компоненты резильентности функционируют по-разному на разных этапах и что аспекту развития должно уделяться основное внимание в концепции резильентности.
Измененная реакция на стресс опосредуется негативным воспитанием
Эффекты материнской депрессии на стрессовую реакцию у детей были разнообразными, меняясь в зависимости от стадии развития, используемых методик оценки и показателей резильентности. В 9 месяцев у младенцев матерей, страдающих депрессией, наблюдалась повышенная реактивность кортизола при воздействии социального стрессора и замедление его нормализации229. В 6 лет сохранялось влияние материнской депрессии на вариабельность кортизола, но только среди детей, которые воспитывались в напряженных, угрожающих и негативных условиях228. Эти результаты подчеркивают важность компонента пластичности/гибкости для стрессоустойчивости. В 10 лет только дети матерей с депрессией, которые воспитывались в негативных условиях, имели более высокий уровень кортизола, такая чрезмерная активация оси гипоталамус-гипофиз-надпочечники (ГГН) опосредовала влияние депрессии на развитие психопатологии221.
Мы также измеряли IgA в слюне, биомаркер иммунной системы, и обнаружили более высокие показатели у детей матерей, страдающих депрессией, в позднем детстве, что указывает на больший стресс. Однако это было выявлено только среди детей, чья синхронность с матерью была минимальна, что свидетельствует о влиянии синхронности на стрессоустойчивость и иммунную систему.
Отцы повышают резильентность
В условиях минимальной синхронности, обеспечиваемой матерью в депрессии, хорошо синхронизированные отношения с отцом выполняли важную функцию в формировании резильентности. Если отцы относились к детям внимательно и отзывчиво, риск развития психических расстройств заметно снижался230. Похоже, что одним из механизмов, с помощью которого заботливое отцовство улучшает резильентность, является изменение семейной атмосферы, что делает семейные взаимодействия более согласованными, гармоничными и вовлеченными, даже когда матери находятся в депрессии231. Эти результаты согласуются с механизмом «социальной моногамии», описанным выше, и предполагают, что связь с матерью открывает ребенка другим аффилиативным связям в семье и обеспечивает резильентность.
В другом исследовании мы наблюдали за родителями и их первенцами в израильском и палестинском обществе от младенчества до дошкольного возраста. Мы обнаружили, что материнская депрессия оказывает менее токсичное влияние на психику и символические компетенции ребенка в палестинском обществе, что было связано с условиями жизни в расширенной семье в этой культуре, которая предоставляла детям широкие возможности для синхронного взаимодействия с другими взрослыми родственниками232.
Окситоцин способствует резильентности
Через 6 и 10 лет у матерей с депрессией и их детей был снижен уровень окситоцина как в слюне226, так и в моче233. Как у матерей, так и у детей распространенность генотипа GG по гену OXTR (rs2254298) была выше, что связано с большей уязвимостью для психических расстройств234. Если у матери имелся аллель А в гене OXTR, вероятность установления диагноза по оси I у ребенка в 6 лет снижалась вдвое226. В 10 лет, если уровень окситоцина в слюне у детей был высоким, влияние материнской депрессии на экстернализированные и интернализированные симптомы ребенка было меньше227.
Аффилиативный мозг подростков
В раннем подростковом возрасте мы оценивали нервную эмпатическую реакцию детей на боль других и привязанность с помощью МЭГ. У детей матерей, страдающих депрессией, мы обнаружили нарушения нервного эмпатического ответа в верхней височной борозде, которая является центром социального мозга, проявившиеся в снижении альфа-активации и более быстром прерывании нервного ответа примерно через 900–1100 мс после стимула. Такое прерывание ответа сочеталось с интрузивным и негативным воспитанием детей и недостаточной синхронностью, которую эти подростки испытывали в младенчестве, что усугубляло пагубные последствия влияния материнской депрессии для развития мозга219.
Для нейробиологической оценки привязанности мы использовали типичный тест – сравнение реакции детей на демонстрацию их взаимодействия с матерью на более ранней стадии и взаимодействия с незнакомцами. Типичная нейронная активация на сигналы привязанности включала мультиритмический альфа-, бета- и гамма-ответ, проявившийся в альфа-супрессии в задней области, и бета- и гамма-активации в обширном кластере справа, включавшем верхнюю височную борозду, веретенообразную извилину и островок. Тем не менее только те дети матерей с депрессией, у кого развилось аффективное расстройство, демонстрировали аберрантный ответ, включающий сниженную реакцию на социальные сигналы и ослабление дифференциации между стимулами привязанности и непривязанности. Эти нарушения предопределялись снижением активности системы окситоцина и снижением синхронности между матерью и ребенком в детстве.
Хотя эти данные указывают на риск нарушений последующих привязанностей у детей матерей, страдающих депрессией, они также показывают, что некоторые дети, растущие в условиях хронической материнской депрессии, более устойчивы, и что нейробиологические компоненты аффилиации являются маркерами резильентности.
Стресс и психотравма в раннем детстве
Когорта переживших стресс и психотравму в раннем возрасте включала детей и их матерей, живущих в зоне непрерывных боевых действий, которые подвергались повторным непредсказуемым ракетным обстрелам в течение почти 20 лет. Обследование детей проводилось в младенчестве, среднем детстве (5–7 лет) и позднем детстве (10 лет), в раннем подростковом возрасте выполнялось нейровизуализационное исследование социального мозга.
Коморбидные психические расстройства у детей с хронической травмой, полученной в раннем возрасте
У детей, растущих в таких травмирующих и непредсказуемых условиях, выявлено увеличение распространенности психических расстройств по оси I в 3–4 раза и заметное усиление интернализированных и экстернализированных симптомов. В сравнении с когортой детей, чьи матери находились в депрессии, особенность этой когорты заключалась в том, что две трети детей имели более одного диагноза, причем у некоторых было три или даже четыре психических расстройства, что свидетельствует о том, что травма ведет к множественным дисфункциям по всему психопатологическому спектру235.
В ходе анализа резильентности в течение первого десятилетия жизни у детей, проживающих в травматичных условиях, мы обнаружили, что матери детей, у которых не было выявлено психических расстройств или отмечалось выздоровление, демонстрировали меньшую выраженность симптомов и проявляли большую синхронность. Такие дети проявляли большую социальную компетентность в позднем детстве (10 лет)236.
Окситоцин как буфер стресса
В исследуемой группе активность окситоцина была связана с резильентностью при травме. Большая активность гена рецептора окситоцина у детей, матерей и отцов отличала детей, у которых развился хроническое посттравматическое стрессовое расстройство (ПТСР), от детей, у которых диагноз был снят в среднем детстве235.
В возрасте 10 лет, в отличие от детей матерей, страдающих депрессией, групповых различий по уровню окситоцина не было выявлено, что свидетельствует о том, что не все дети, растущие в зоне военных действий, демонстрируют фундаментальные биологические нарушения аффилиации и что некоторые матери способны, прилагая значительные усилия, смягчить опасные последствия войны для своего ребенка. Однако уровень окситоцина у матерей, живущих в условиях войны, был ниже, что свидетельствовало об огромном напряжении при воспитании ребенка в травматичных и непредсказуемых условиях. Множество таких изменений затрагивают эндокринную и нервную систему матери.
Выявлена эндокринная синхронность между матерью и ребенком. Когда уровень материнского окситоцина был низким и снижался уровень синхронности, у детей проявлялось значительно больше симптомов. Но этого не наблюдалось в случаях, когда у матери сохранялся высокий уровень окситоцина и стиль воспитания был заботливым и неинтрузивным120.
Реакция на стресс
Мы измеряли базальный уровень кортизола и его колебания у матери и ребенка в раннем детстве, в позднем детстве и в раннем подростковом возрасте. Уровень гормонов оценивался в волосах и слюне. В раннем детстве уровень кортизола и альфа-амилазы слюны, маркера симпатической реакции на стресс, различались у детей с ПТСР и без него. У детей без ПТСР уровень гормонов был значительно выше, в то время как у детей с ПТСР отмечались более низкие уровни236. Эти данные свидетельствуют о том, что в условиях хронической травмы в раннем детстве большая активация оси ГГН способствует резильентности, а не увеличивает риски.
Через 10 лет базальные показатели и колебания гормонов оси ГГН были повышены только у детей, проживавших в условиях военных действий, у которых развились психические расстройства, и это были дети матерей с более выраженной активацией оси ГГН и меньшей синхронностью в детско-родительских отношениях73. Мы предполагаем, что «матери стояли между войной и ребенком», и в случаях, когда мать способна совладать с собственным стрессом и защищать ребенка от внешней травмы, она способна сдерживать стрессовую реакцию ребенка.
Тем не менее в раннем подростковом возрасте дети и их матери в целом демонстрировали более высокие и менее вариабельные уровни кортизола, что позволяет предположить, что хроническое воздействие непредсказуемого стресса само по себе является фактором риска, независимо от взаимоотношений. Возможно, такая уязвимость сопровождает ключевые периоды развития, в частности переход к подростковому возрасту237. Показатели иммунных биомаркеров были выше у матерей и детей, проживающих в зоне боевых действий, что подчеркивает значительное истощение иммунной системы в условиях хронического стресса и согласуется с моделями аллостатической нагрузки и реакции на стресс238.
Мозг ребенка и матери
В этой когорте, в отличие от двух других, мы рассматривали мозг матери и ребенка в идентичных условиях, пытаясь оценить, как хронический стресс воздействует на нервные системы обеих сторон диады. Мы обнаружили, что изменения в функционировании мозга были опосредованы историей взаимоотношений, что привело к различному (благополучному и неблагополучному) созреванию социального мозга у детей.
Мы оценивали коннективность и мощность сети пассивного режима работы головного мозга (СПРРГМ, default mode network – DMN), обеспечивающей чувство самосознания, переключения внутреннего и внешнего внимания и автобиографическую память239-241. И у матерей, и у детей были обнаружены и нарушения коннективности СПРРГМ, и ее мощности, что еще раз подчеркивает роль пластичности в формировании резильентности и указывает на недостаточную способность отдельных структур объединяться в единую систему, которая обеспечивает основу самосознания.
У матери обнаруженные нарушения СПРРГМ происходили на уровне альфа-ритма, основного ритма бодрствующего зрелого мозга, тогда как нарушения СПРРГМ у детей происходили в тета-диапазоне, биомаркере развивающегося мозга242. У детей с ПТСР наибольшие нарушения затрагивали тета-соединения. Нарушение паттернов тета-связности было опосредовано материнским интрузивным, провоцирующим тревогу воспитанием и более высоким уровнем кортизола в более позднем детстве, что подчеркивает долгосрочные негативные последствия непредсказуемого воспитания, при котором родитель не управляет своими эмоциями, для ключевой нейронной системы243.
У детей не было обнаружено никаких групповых различий нейронального эмпатического ответа на страдания других. Этот ответ приводил к альфа-активации большого кластера, включавшего дополнительную моторную область, часть нейросети, отвечающей за самосознание, и срединную поясную извилину, узел СПРРГМ. Синхронность, которая была снижена в когорте живущих в условиях боевых действий, опосредовала влияние ранней травмы на нервный эмпатический ответ, и дети, получившие больше синхронности в детстве, демонстрировали более активную реакцию на страдания других244. Нервная эмпатическая реакция матерей также имела нарушения, но они были специфическими для мозга взрослого245.
Недоношенность
Наш проект с использованием «метода Кенгуру» является единственным существующим исследованием, в котором оценены эффекты разлуки с матерью, отсутствие структурированного контакта с ней на созревание функций жизнеобеспечения у младенцев в долгосрочной перспективе. Матери недоношенных детей с низкой массой тела при рождении (<1750 г) были рандомизированы в две группы. В одной применялась экспериментальная методика (контакт «кожа к коже» не менее часа в день не менее 14 дней подряд в течение периода выхаживания), в другой – выхаживание младенца осуществлялось стандартным способом в инкубаторе.
Диады обследовали семь раз в течение первого десятилетия (до вмешательства, при выписке, в возрасте 3, 6, 12 и 24 месяцев, в 5 и 10 лет). По достижении детьми молодого возраста (18–20 лет) мы посещали их на дому, чтобы оценить их отношения с матерями, гормональные показатели и исполнительские функции, а в течение следующего месяца проводили функциональную магнитно-резонансную томографию.
Мы обнаружили, что обеспечение телесного контакта с матерью у людей влияло на те же системы, что и у других млекопитающих. «Метод Кенгуру» улучшал у новорожденных функционирование вегетативной системы, организацию циклов сна–бодрствования, ориентацию и обработку информации. В то же время он обеспечивал потребность в материнской заботе в период новорожденности246.
В соответствии с нашей моделью поэтапного развития регуляторных функций138 эти улучшения в физиологической регуляции и взаимодействии с матерью способствовали улучшению резильентности и влияли на динамику развития. Новорожденные демонстрировали лучшую модуляцию возбуждения при обработке сильных возбуждающих стимулов в 3 месяца, лучшее исследовательское поведение в 6 месяцев и лучшие способности к самоконтролю в 1 и
2 года. Умственные, но не моторные способности были лучше в экспериментальной группе через 6, 12 и 24 месяца247,248. Также улучшалась синхронность между матерью и младенцем, а у матерей увеличивалось количество грудного молока в ответ на увеличение уровня окситоцина249. После использования «метода Кенгуру» более высокая синхронность отмечалась при каждом обследовании в первые годы, и большая взаимность при социальных взаимодействиях была связана с лучшими когнитивными и регуляторными способностями210.
Через 10 лет мы обнаружили, что лучшие регуляторные способности субъектов из экспериментальной группы сохранялись. Мы обнаружили более высокую дыхательную синусовую аритмию и ее большую чувствительность к эмоциональному стрессу, что указывает на более адаптивное функционирование вегетативной нервной системы. Оценка сна выполнялась с помощью актиграфии в течение пяти ночей подряд, и у детей, к которым был применен «метод Кенгуру», организация сна была лучше, периоды бодрствования короче. Кроме того, реакция оси ГГН у детей из экспериментальной группы на социальный стрессор вызывала сниженный кортизольный ответ на стресс и отмечалось более быстрое восстановление250. Что касается когнитивных способностей, к 5 годам больше не было различий общего IQ, но дети, у которых применялся «метод Кенгуру», имели лучшие исполнительские способности, рабочую память и когнитивную гибкость через 5 и 10 лет.
В целом наши результаты позволяют выделить системы, на которые влияют компоненты устойчивости, связанные с материнским телом и адаптивными формами ухода, и подтверждают наличие их связей с совладанием со стрессом, гибкой реакцией на условия окружающей среды, модуляцией возбуждения и внимания, а также способностью участвовать в взаимном диалоге.
В молодом возрасте для оценки эмпатической реакции мозга на эмоции других в группе детей, у которых применялся «метод Кенгуру», и контрольной группе изучали «эмпатическую точность», важную детерминанту эмпатической реакции251,252 и дифференцированные реакции на стресс, печаль и радость других людей. Используя комплексный анализ, мы обнаружили три структуры, которые показали очень разную активность в ответ на эмоции: миндалина, передняя островковая доля и височный полюс. Уровень синхронности, прослеженный в динамике от младенчества до юности, опосредовал участие в групповых взаимодействиях и гибкий эмпатический отклик социального мозга на эмоции других. Таким образом, увеличение синхронности благодаря «методу Кенгуру» позволило ранним переживаниям привязанности сформировать гибкую нервную реакцию на эмоциональные состояния других.
ВЫВОДЫ
Резильентность является основным понятием клинической науки и практики, которая до сих пор не имеет исчерпывающей биоповеденческой концепции. Два основных упущения в современных моделях резильентности связаны с ее рассмотрением исключительно в рамках нейробиологии страха и недостаточным вниманием к ее стадийному развитию. Более того, большинство моделей дают резильентности отрицательное определение (то есть отсутствие симптомов после травмы), в действительности не давая определения, что такое, собственно, резильентность.
Мы утверждаем, что для понимания резильентности необходимо принять во внимание особенности первоначального состояния млекопитающих. Детеныши млекопитающих рождаются с двумя важными ограничениями: при рождении их мозг незрел, и они поддерживают тесную близость с кормящей матерью. Таким образом, все системы, обеспечивающие резильентность, стрессоустойчивость, адаптацию и выносливость, у млекопитающих созревают под влиянием условий, которые обеспечиваются физиологией материнского тела и родительским поведением.
Мы предлагаем модель резильентности, основанную на нейробиологии аффилиации, развивающейся научной области, которая описывает нейронные, эндокринные, генетические и молекулярные процессы, лежащие в основе нашей способности формировать связи, любить, заботиться, сопереживать и принадлежать к социальным группам.
Наша модель выделяет три основных нейробиологических компонента аффилиации, которые обеспечивают резильентность. К ним относятся окситоциновая система, аффилиативный мозг и биоповеденческая синхронность.
Окситоциновая система, участвующая в процессах пластичности на клеточном, молекулярном уровнях и уровне нейрональных сетей, обеспечивает формирование привязанности, способность млекопитающих справляться с трудностями с помощью отношений и влияет на иммунную систему.
Аффилиативный мозг человека, имеющий происхождение от материнского мозга грызунов, расширился за счет включения структур более высокого порядка, которые обеспечивают эмпатию, подражание и ментализацию, и служит базой для развития других аффилиативных связей, включающих романтическую привязанность, близкую дружбу и наставничество. Он чрезвычайно пластичен, обеспечивает передачу от одного поколения к другому в ранние сензитивные периоды и формирует социально-эмоциональные компетенции.
Биоповеденческая синхронность представляет собой координацию биологических и поведенческих процессов во время социального взаимодействия, посредством этого механизма зрелый мозг матери внешне регулирует незрелый мозг ребенка и настраивает его на социальную жизнь. Биоповеденческая синхронность человека опирается на механизмы, с помощью которых у животных согласованное социальное поведение способствует разнообразию и адаптации в процессе эволюции, и развивается в рамках детско-материнской связи на основе биологических ритмов плода еще внутриутробно, на базе которых мать строит социальный невербальный «танец» в течение первых месяцев жизни. Этот синхронный обмен развивается, усложняется, превращаясь в диалог, внутри которого есть взаимность, близость и признание множества точек зрения, распространяется от взаимоотношений матери и ребенка на другие привязанности и существует на протяжении всей жизни, намечая ключевую траекторию в развитии резильентности.
Наша модель предлагает три принципа резильентности.
К ним относятся пластичность, социальность и осмысленность. В то время как первые два являются общими у человека с животными, последний специфичен для человека. Все три принципа обеспечиваются окситоциновой системой, аффилиативным мозгом и биоповеденческой синхронностью благодаря их участию в нейрональной и поведенческой пластичности, в формировании привязанности и социальности. Они также обеспечивают способность придавать смысл травме через культурные и духовные системы и аффилиативные акты, которые выходят за рамки личности.
Эта модель подтверждается лонгитудинальным исследованием трех когорт, в которых испытуемых обследовали в период от рождения/младенчества до юности. Каждая когорта соответствовала одному из типов нарушений детско-материнских отношений: связанных с матерью, ребенком или условиями среды (материнская депрессия, преждевременные роды и хроническая травма в связи с проживанием в районе военных действий), – которые оказывают долгосрочное влияние на мозг ребенка, его поведение и благополучие. В каждой когорте гипотезы были построены на конкретных исследованиях животных моделей, описывающий «недостающий компонент» (симпатия и груминг, изменчивая доступность кормовой базы и материнской близости). Мы неоднократно оценивали психическое состояние, взаимоотношения с родителями, синхронность, уровень окситоцина и гормонов стресса, когнитивные и регуляторные функции, с целью установить факторы, которые определяют резильентность у детей. В подростковом возрасте или в ранней молодости проводилось нейровизуализационное исследование социального мозга.
Нарушения в развитии возникали в разных условиях; тем не менее, они были предсказуемыми и в основном проявлялись во взаимодействиях с людьми, при этом некоторые дети демонстрировали значительную резильентность. Нейробиологические компоненты аффилиации – синхронность и система окситоцина – показали себя как факторы, способствующие резильентности в процессе развития. Эндокринная синхронность (гормональная согласованность между показателями окситоцина и гормонами стресса у матери и ребенка) проявила себя как фактор, определяющий риск дезадаптации, что свидетельствует о постоянном биологическом воздействии матери как внешнего фактора, регулирующего резильентность. В позднем детстве социальные навыки детей, обеспеченные синхронностью, выполняли роль важных показателей резильентности. Созревание регуляторных функций происходило на базе друг друга, и более высокий уровень регуляции улучшал последующее функционирование. Особое значение имели интервенции во время ранних сензитивных периодов, как, например, в результате обеспечения контакта «кожа к коже» матери с младенцем у недоношенных детей.
При нейровизуализационном исследовании социального мозга мы обнаружили изменения, определяющие резильентность. В то время как у детей, воспитанных матерями с хронической депрессией, отмечалось прерывание эмпатического ответа нервной системы, не у всех детей, растущих в травматических условиях, имелись нарушения, а только у тех, у кого синхронность с матерью была недостаточной. Нейробиологическая основа привязанности была нарушена у детей матерей, страдающих депрессией, но только у тех, у кого в последующем развилось аффективное расстройство. Аналогичным образом при нейробиологической оценке эмпатической точности недоношенные дети, имевшие достаточную синхронность с матерью, показали адекватный социальный нейронный ответ.
Таким образом, опираясь на философские и нейронаучные модели 20-го века, которые формулировали конкретный, поведенческий подход к познанию и поведению и размыли различие между мозгом и разумом, наша модель имеет целью направить внимание на системы, которые поддерживают нашу способность формировать аффилиативные связи, вступать в социальные группы и использовать отношения, чтобы справляться со стрессом, как специфические способности человека для того, чтобы противостоять, и даже процветать, перед лицом травмирующих обстоятельств.